The
Skull Structure of the Mosasaurus,
explained by means of a description of a new species
of this genus
by
Dr. AUGUST GOLDFUSS,
DIRECTOR OF THE ACADEMY
_______________________
WITH FOUR LITHOGRAPHS
Communicated to the Geological Section of the Natural
Scientists’ Conference in Mainz, September 1842
(ACTA OF THE IMPERIAL LEOPOLD CHARLES ACADEMY OF NATURAL
CURIOSITIES, VOL. XXI, P. 1)
At a conference of the Natural Scientists in our
city I have already had the honor of displaying to the Geological Section the fragments of
a fossil skeleton, encased in stone, which His Highness Prince Maximilian zu Wied
had presented to our museum as a highly valued gift. They had been discovered by Major
O’Fallon, former Agent of the Indian Nations of the Upper Missouri, between Ft.
Lookout and Ft. Pierre.*) They were brought to St. Louis and placed in the garden of their
owner at that time.**) The bones were encased in a very hard fine-grained, dense limestone
several inches thick, which in the interior of the larger pieces is bluish gray; closer to
the exterior it is ocher yellow and softer, and on the surface black and bituminous. Where
the stone covering of the bones is thin, it is completely black and bituminous. The
skeleton was buried very slightly below the slope edge of the mountain stratum, whose
smooth, partly polished surface could be distinguished by the fragments of the opposing
fracture-surface. Here and there parts of the skull and vertebral processes projected out
from the rock and were slightly weathered, and partly broken off, and coated with coal
dust.
____________________________
*) The Journey of Prince Maximilian zu Wied to the Interior of North
America. I. p. 348.
**) Loc. cit., II, p. 78.
4 (176)
A. Goldfuss
The bones, completely encased in stone, are impregnated with fossilization matter,
dense, hard and brittle. The stone block, over two feet long and thick, which contains the
skull, had been broken into several pieces, whose undamaged fracture surfaces could be
fitted together, however, and the vertebrae of the extended vertebral column,
approximately 12 feet long, were in several cylindrical pieces. In the larger pieces there
are very many crushed shells of larger and smaller species of the genus Inoceramus.
The smaller ones can be identified as Inoceramus Barabini, and In.
alveatus Morton *); the larger ones are large shells of Inoceramus Cuvieri.
Since large Baculites **) are also found in those mountain ranges, there is
no doubt that the rock belongs to the Cretaceous formation . This extends, as
Morton ***) remarks, between the Mississippi and the Rocky Mountains; it forms the hills
on the banks of the Upper Missouri, §) which resemble towers and ruins, similar to those
in the Elbe valley near Dresden, and it contains a quantity of large gypsum crystals §§)
as in Silesia. On top of it is a layer of bituminous brown coal, which can be seen
extending several hundred English miles in the hills along that river (the
Missouri).§§§) The bituminous character and the coal dust clinging to the projecting
parts of our skeleton allow the assumption that it was covered by the coal stratum.
In the immediate area of the bone the stony covering is so
hard that small pieces
_______________________________________
*) G. Morton. Synopsis of the organic remains of the Cretaceous
Group of the United States, page 62, tab. 13, fig. 11; tab. 17, fig. 3 and 4.
**) Prince M. von Wied’s Journey, I, pp. 331-349.
***) Morton, loc. cit., p. 25
§) Prince v. Wied Journey. I, pp. 331-349.
§§) Loc. cit., p. 333.
§§§) Loc. cit., p. 324, 334, 348
Skull Structure of the Mosasaurus
(177) 5
could only be chipped off with a steel mallet. Since moreover the bones are much more
brittle, no force could be applied during their uncovering, and therefore this difficult
labor achieved the desired result only after a number of years. It was ultimately possible
to disclose the skull in a reasonably complete form.
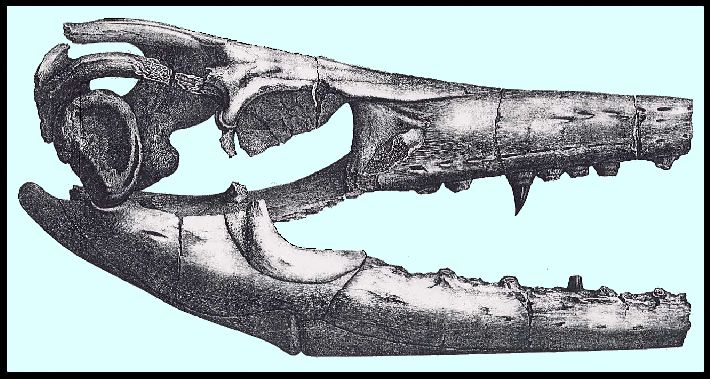
Only the tip of the snout and the zygomatic arches are missing, as well as the temporal
bone, and the tympanic bone and the mastoid bone of one side. The two rami of the lower
jaw could not be completely freed from the stony covering, and its chin symphysis is
likewise not present. The vertebrae of the spinal column lie mostly in a continuous row
next to one another; however, the processes protruding outside the stony cover were more
or less shattered, or they crumbled at the attempt to free them from the stone. Here and
there short pieces of ribs were found. Only a few small fragments of the extremities, the
pelvis and the shoulder-blade were present.
Since the pterygoid bones were not covered by stone, it could be conjectured from the
very beginning that this skeleton belonged to the genus Mosasaurus. This
conjecture became a certainty during the preparation of the skull. Its structure fills in
the anatomical features of the Mosasaur which the Maastricht fragments *) did not provide,
and it shows that it belongs to a different species than the Mosasaurus Hofmanni,
differing not only in its smaller size, but also in some other features. –The
complete ossification of all parts as well as the frequently observed filling out of the
teeth prove that the specimen had reached its full development with only half the length
of Mosasaurus Hofmanni.**)
____________________________
*) Cuvier, Oss. foss., page 310, tab. 18, 19 20.
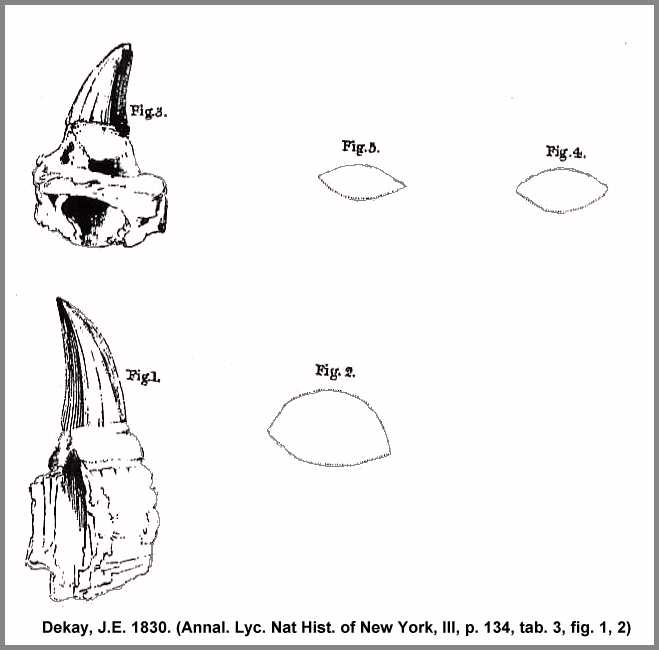
**) It can not be determined whether the dental remains from the Cretaceous of
New Jersey as illustrated by Dekay (Annal. Lyc. Nat Hist.
of New York, III, p. 134, tab. 3, fig. 1, 2) differ from those of the Maastricht
specimen or not, based on its size, which seems to be predominant, since our museum owns a
tooth from St. Petersburg, which is also 5 inches in length, including the socket.
6 (178)
A. Goldfuss
Since the end of the snout of both the Maastricht skull and our own has been lost, no
specific difference can be determined by the number of jaw teeth. In both eleven teeth
remain on each side of the maxilla. In the complete lower jaw of Mosasaurus Hofmanni
there are 14 teeth; in each of the broken-off rami of the mandible of the American
specimen there are only eleven. In both specimens the tooth crowns, which are covered by a
brown shiny enamel, are seated on a dental capsule which has been modified into an
ossified base which has grown into the alveoli. Some of these capsules are hollow on the
inside, and some are filled up. The bases and their alveoli extend down over 2/3 of the
total height of the walls of the jawbone, and the replacement teeth are breaking through
laterally in the interstices. Likewise the tooth crowns of both species are slightly
compressed laterally (Plate IX, Fig. 4), and at the tips
slightly curved towards the back and divided into two halves by a sharp straight ridge (Plate IX, Fig. 6), which is cross-striped and finely
serrated, the inner part somewhat larger than the outer part. Its surface is polygonal, as
on the outside five and on the inside seven pyramid-shaped surfaces of unequal size can be
seen, which are already taking shape in the small replacement teeth.
On the other hand the pterygoid bone has two distinctive features. In the Maastricht
skull its tooth edge clearly curves downward towards the front and has only 8 teeth. In
our skull, however, this edge is not perceptibly curved, and it holds 10 teeth.
Skull Structure of the Mosasaurus
(179) 7
These features thus allow us to view the American specimen as a distinctly different
species, to which I wish to give the name
Mosasaurus Maximiliani
in honor of Prince Maximilian zu Wied, a scientist, well known for his travels to
Brazil and North America, who rescued these remains from destruction and who transported
these heavy masses of stone thousands of miles to us here, in order to present them to a
German museum for the purpose of scientific study.
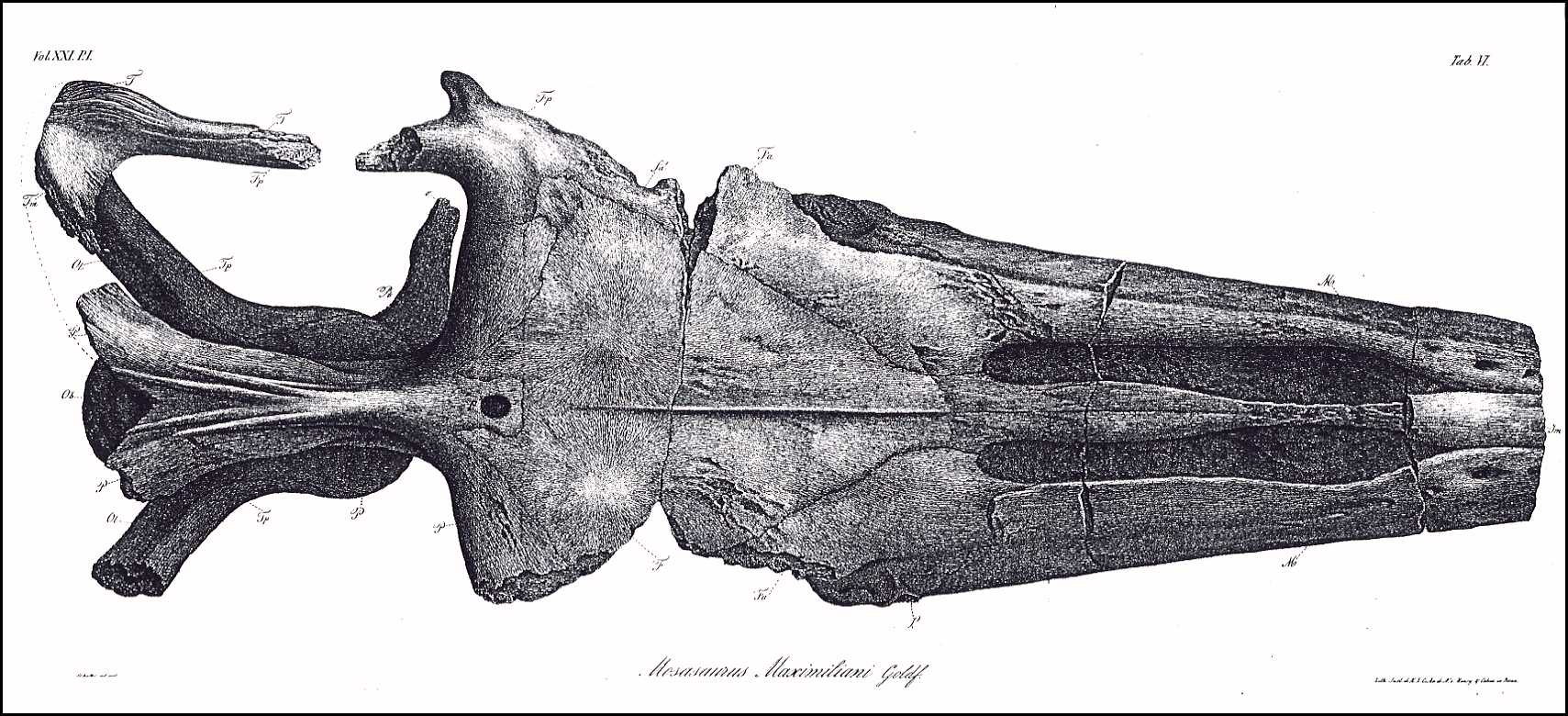
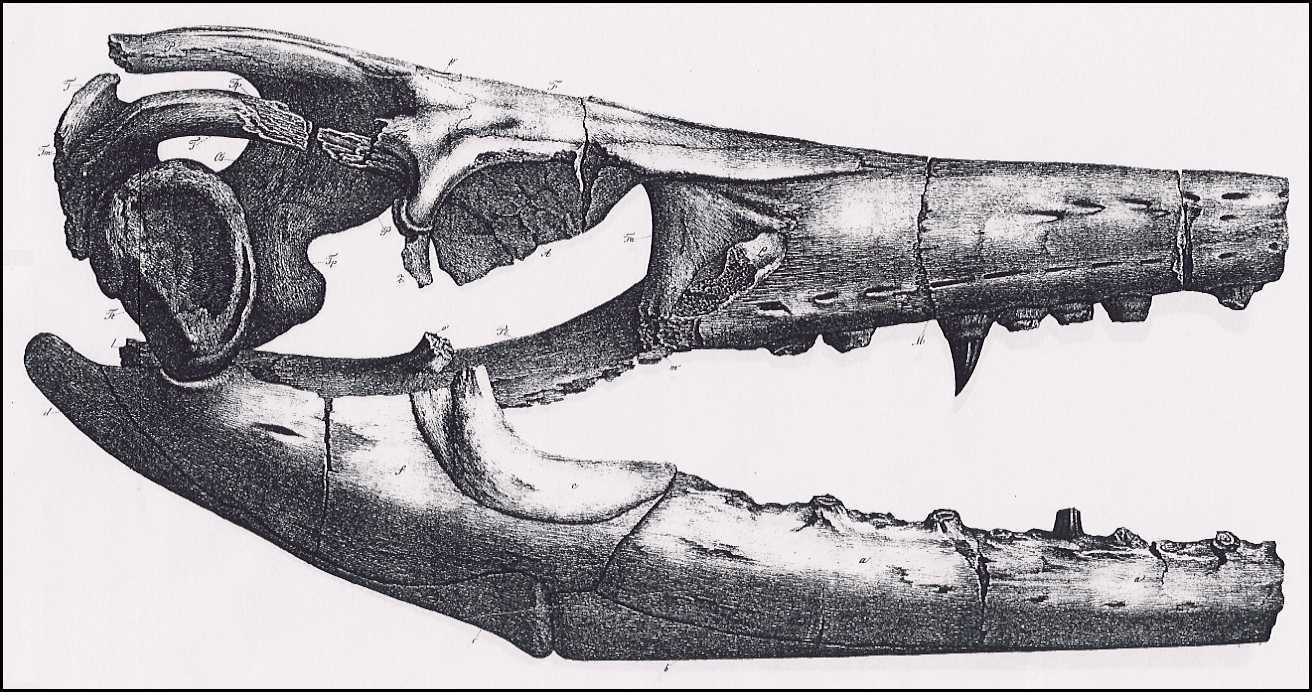
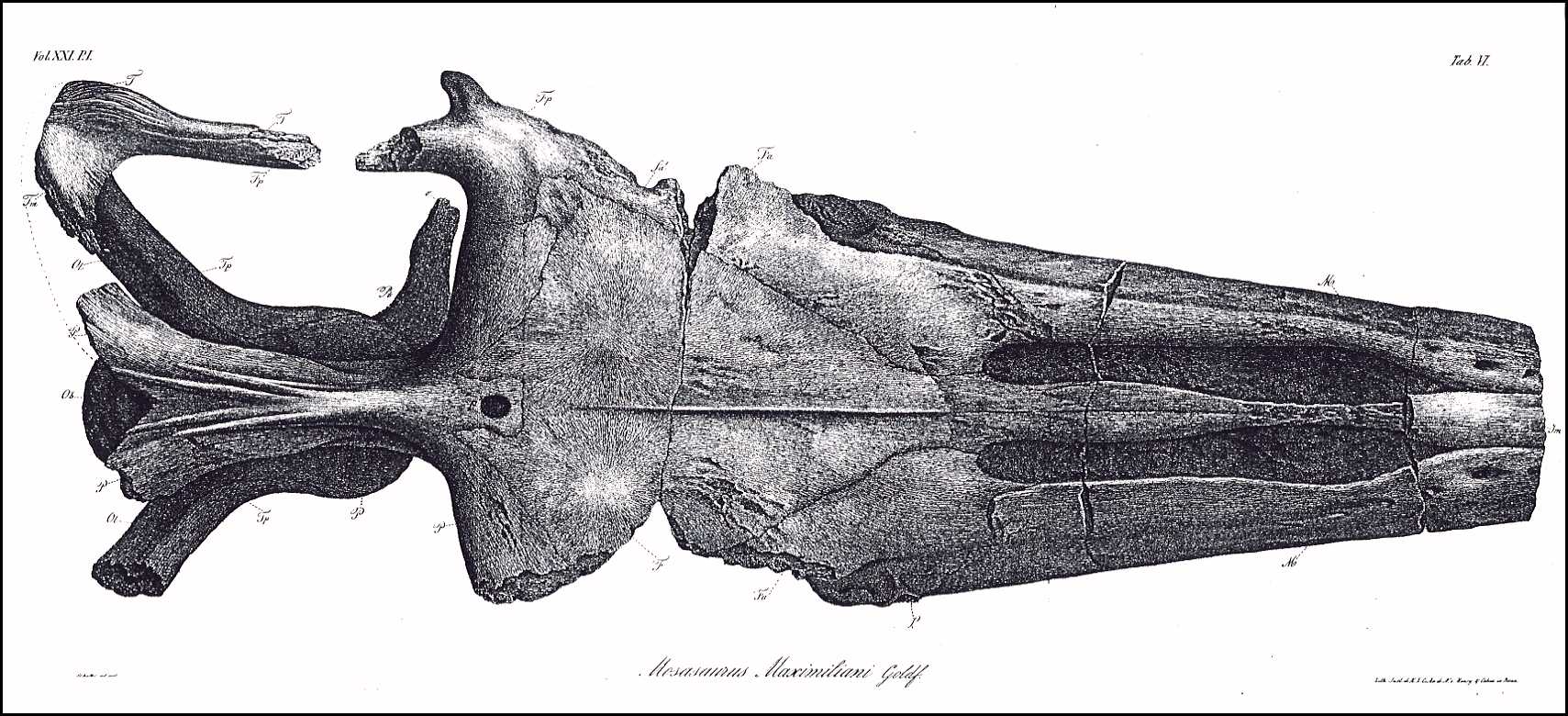
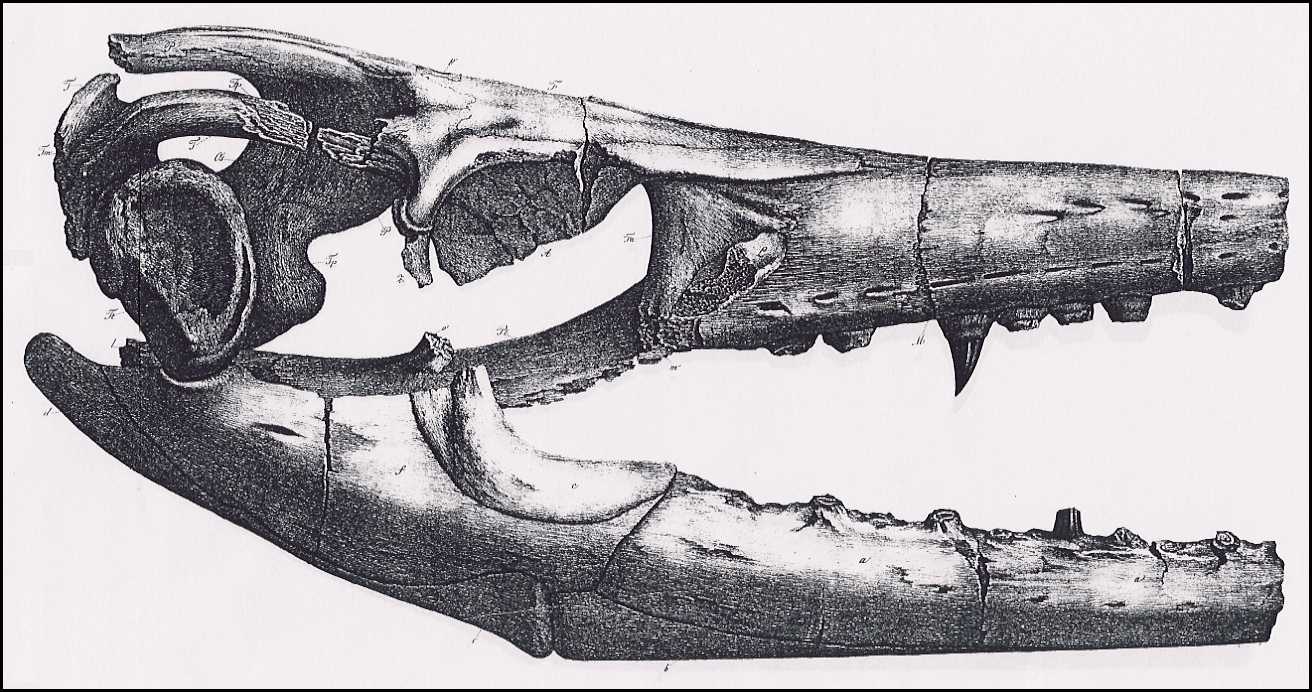
The appended drawings (Plates VI, VII, VIII) depict the
skull in its natural size, following precise measurements; therefore, the specific
indications of its dimensions are unnecessary, and are also superfluous because the
missing nose tip prevents the determination of the total length and comparison with other
skulls. But even lacking the determination of precise relative dimensions, this skull is
still complete enough to provide the knowledge of its genus.
The low, elongated shape of the front of the head, the narrow, long nostrils, the
formation of the mandible and the presence of palatal teeth confirm Cuvier’s
statement, that this genus of animal must be assigned to a place in the classification
between the Monitors and the Iguanas. But if we proceed to
examine the individual parts of the skull structure, we are surprised to find here a
common center in which are combined not only the properties of the two genera mentioned,
but also of most other saurians; however, a number of features remain which belong to this
species alone and distinguish it from all others.
8 (180)
A. Goldfuss
It is to be supposed that the tip of the snout shares similarities with that of the Monitor,
that it therefore was broken off shortly before the widening of the intermaxillary bone.
One can judge the size of the narrow nasal process of this bone by the distance between
the nostrils (Tab. VIII. Jm.).
At the back it is so closely grown together with the adjacent simple nasal bone (N.)
that one can not recognize its boundary on the somewhat damaged surface. It was simple,
not doubled, as A. Camper had supposed.
The narrow nasal bone (N.) is connected with the
medio-frontal bone by three deeply cut prongs. The long nostrils are similar to those of
the Monitor, but narrower by half and somewhat shorter. At the anterior end
they dwindle into a narrow furrow; in the first third of their length they widen, then
become a bit narrower, and their posterior end is rounded, while they are pointed in the Monitor.
There is no trace of the spoon-shaped opercular bones, which in the Monitor
close off the front half of the nasal cavities.
The maxilla (M.) in its relative length resembles that of the
Monitor, but, like the skull as a whole, it is considerably narrower,
somewhat higher, with no depression in front of the nostrils, but rather, it rises evenly
in a somewhat inclined plane. In its upper third, parallel to the nostrils, there is a row
of 6 holes whose oval openings are directed towards the back and turn downwards. These
holes are lacking in the skull of the Monitor. A second row of 11 holes,
whose oval openings turn forward and downward, runs parallel to the edge of the teeth,
some distance away, as in the Monitor, but at more regular intervals.
Between the two rows, at the front, in front of the nostrils down to the broken surface of
the nose tip, there are three more similar holes, whose
Skull Structure of the Mosasaurus
(181) 9
openings also turn frontward. In Cuvier’s drawing of the broken maxilla (Tab. 18,
Fig. 1.h.) only one of the holes of the lower row is indicated, but on the plaster cast of
the skull some of the upper row can be detected. The tooth edges are one inch wide, and
are bordered inside and outside by a furrow. The oval bases of the teeth press up against
one another at their inferior end. The front two were split longitudinally by the
fracture, and reveal that the jaw bone is penetrated through 2/3 of its height by the
tooth holes. These appear to be the front-most teeth of the maxilla. The zygomatic process
of this bone (m.) was broken off along with the zygomatic bone,
and it was so low that it could not contain any tooth sockets, as in the Monitor,
and could serve as a base for only a narrow zygomatic bone.
The medio-frontal bone (F.) does not consist of two bones, as
is the case in the Monitor, and its rear edge is not straight, as in other
lizards, but it is a simple, smooth, inverted delta-shaped bone, whose anterior and
posterior points are cut off. In its center line it has a low ridge which quickly flattens
out toward the back. Its long, almost straight edges converge towards the serrated portion
of the tip, which adjoins the nasal bones. The blunt point, which would be formed by the
converging shorter posterior side edges, is carved out in a semicircle, and this cut-out
is also shown in A. Camper’s drawing of the inner surface of a frontal bone from
Maastricht (Oss. foss. tab. 20, fig. 1). At the anterior end one can make out a
fine crack, the trace of a separation into two halves which existed in a juvenile stage.
Also, fibers of this bone radiate towards the back from two points on the sides.
The anterior frontal bone (Fa.) is very large and joins the
posterior frontal bone
10 (182)
A. Goldfuss
(Fp.) by means of a long process, so that the two encircle
the medio-frontal bone (F.) and form the rim of the orbit. A
small cut out (fa.) near its posterior end is an indication of
the hole which in the Chamaeleo bifurcus is found between the process of the
anterior frontal bone and the medio-frontal bone. As the anterior third of the orbital
rim, it forms a projecting edging, descends vertically at a right angle, at its lower edge
it makes some large creases in order to approach the lachrymal bone, and bends with a
curve to form the anterior wall of the orbit.
The lachrymal bone (L.), which lies between the lower edge of
the anterior frontal bone and the zygomatic process of the maxilla, is very narrow, a
longish rectangle of a small size, and it is broken off above the tear duct. It descends
steeply from front to back, forms a bumpy surface, and at its inner edge it has a cut-out,
bordering on the large opening which is formed by the adjoining bulge of the anterior
frontal bone.
The parietal bones (P.), as in most saurians, have fused
together into one piece. Their frontal area is very short, not as long as in the Monitors,
and even shorter than in the Iguanas. It is also narrower than in the
former, because it does not attain the width of the medio-frontal bone, but rather it is
compressed on both sides by the posterior frontal bone. In the middle it extends into the
carved out tip of the medio-frontal bone by means of a spoon-shaped extension, and on the
latter it is pierced by the parietal foramen. This is an arrangement which is found in no
other lizards. In its continuation towards the back, the frontal surfaces become narrower
than in the two genera mentioned here, and at the mid-point of its extent it separates
into the forked mastoid processes, whose upper narrow edge-moldings begin far to the
front. They are narrower than in the Monitor and
Skull Structure of the Mosasaurus
(183) 11
the other lizards, and their exterior surface slants so slightly downward that it is
almost horizontal, and through this peculiarity identifies the genus. The temporal areas
of the parietal bone descend so steeply, like a sloping roof, that the dome of the skull
within the temporal cavity has a lateral diameter only half as long as that of the Monitor;
indeed, it is even smaller than in Istiurus and Iguana, and,
moreover, it narrows towards the back.
The posterior frontal bone (Fp.) abuts the posterior external
corner of the medio-frontal bone and the outer edge of the frontal surface of the parietal
bone. Its zygomatic process is narrow, and there is a small fragment of the zygomatic bone
(Z.) still attached. The temporal process on its outer surface
slants toward the exterior, and as in the Monitor it is received by the
forked opening of the temporal bone (T.) The boundary of this
insertion is indicated on the exterior by a groove.
The existing piece of the zygomatic bone reveals that the zygomatic arch was closed and
very narrow and weak, as is also demonstrated by the broken zygomatic process of the
maxilla. The orbital opening which is enclosed below by the zygomatic bone is oval, twice
as long as it is high, and much lower than in the Lacertia, for which reason
the superciliary bone (os superciliare) is lacking. The large eye was surrounded by
a bony ring whose large plano-convex plates lie in the right orbit, shoved on top of one
another.
The mastoid bones (Tm.), the continuation of the bony arches
of the temporal bone, have their outer surface inclined upwards, as does the temporal
bone, but they assume a vertical direction at their connection with the mastoid process of
the petrosal bone (Tp.).
The temporal bone (T.) differs greatly from that of the Monitor,
for at its posterior end it forms a broad, triangular, somewhat depressed area which is
12 (184)
A. Goldfuss
framed by rough bulges, and with its posterior edge it abuts the mastoid bone. Its
outer tip is broken off, along with the ends of the lateral occipital bone and the mastoid
bone, which converge here.
Since the temporal portion of the parietal bone extends far downwards, the petrosal
bone (Tp.) is shorter than in the Monitor, and
also the lower wing of this bone, which is directed forward to close off the skull cavity,
has a shorter length, but a considerable thickness.
The extraordinary shortening of the dome of the skull, as well as its narrowing, is
also notable. The edge of the superior occipital bone (Os.) is
compressed from both sides to such an extent that it forms an acute-angled roof, whose
peak is shorter and drops off more steeply than in the Monitor.
The body of the basilar bone (Ob.) is narrower and shorter by
one third than in the Monitor. In its relative dimensions it resembles that
of the Istiurus, it is also concave underneath, and has pronounced gnarls
before its anterior end..
The body of the sphenoid bone is covered by the pterygoid bones and appears to have
been partially lost. The occipital foramen is a transverse oval, proportionally smaller
than in the Monitor, and filled with a very hard pebbly stony mass. The
lateral occipital bones (Ol.) show no notable differences from
the structure of the Monitor.
The two pterygoid bones (Pt.) have the same characteristic
form as in the Mosasaurus Hofmanni. They are slender, as in the Monitor,
but they bear teeth like those of the Leguane anolis and Lacertia.
In front they narrow to a point and attach to the inner edge of the palatine bones (Pl.),
and they reach the vomers with their tips.
Skull Structure of the Mosasaurus
(185) 13
For 2/3 of their length they lie close beside one another, and they diverge only behind
their connection with the sphenoid bone. Since they are almost completely preserved in the
Maastricht skull, although out of their natural position, their shape can be clearly
recognized in Cuvier’s drawing (Oss. foss., tab. 18, fig.1, K, l, m). In our
skull their position is also visible, and only the right one was pushed back slightly.
Instead of forming a horizontal surface with their anterior half, they bend lengthwise to
form a groove, so that their inner edges extend down to the palate and the outer edges
stand upright. Their concave lower side faces towards the outside, and the convex upper
side is turned inward. In the median line of the latter they meet. Cuvier took the bone of
the right side, whose outer surface is visible (o', k', l', m'),
to be the one on the left, probably because he viewed the inner surface of the left bone (k,
l, m) facing towards the eye (which lies above the right bone) as the outer
surface. The correct position of these bones is clear from our illustration, Plate IX, Fig. 7. The teeth are on the inner edge which
slopes up and across, and between the two rows of teeth therefore a groove is formed
running from the palate. In Mosasaurus Hofmanni
the dental edge makes a distinct downward curve, which is much flatter in Mosasaurus
Maximiliani. The groove begins in the area of the
anterior edge of the orbit, ends beneath the posterior frontal bone and contains 10 teeth,
with bases almost touching one another, which were not covered by the stony layer, and
whose crowns were therefore lost because of weathering. In the Maastricht skull, a plaster
cast of which was presented to me by the most laudable generosity of the museum in Paris,
these palatal teeth are barely half as large as those of the jaw, with the same curve, but
crowded somewhat more closely together. Such a tooth
14 (186)
A. Goldfuss
in our collection (Plate IX, Fig. 5) also has the
same sharp, finely serrated and cross-striped ridge, but it is, by the way, smooth and not
divided into pyramidal surfaces.
In its posterior half the pterygoid bone divides into three processes, which in our
drawing (Plates VII, VIII)
are given the same letters used by Cuvier. The first process (o.), which points outward,
connects with the transverse bone. It is rounded off and even narrower than in the Monitor,
whereas in the other Lacertia it descends with a fairly wide wing shape. The
middle one (l.) forms the process of the pterygoid bone to the
tympanic bone, and the lower one (m.) is a continuation of the dental edge, and contains
the base for the pterygoid process of the sphenoid bone. The tympanic processes diverge
towards the back; they are compressed from both sides, higher and thinner than in the Monitor,
and their outer surface is directed down and a bit inwards. Only one of these is
preserved; the other is broken off. The lower wing (m.) is
lacking on both sides, so only the fracture surfaces (m'.) can
be seen. In other lizards it forms only a stump below the depression of the sphenoid
process; but in Istiurus pustulatus a slight lengthening shows up.
[Editor's note: Istiturus pustulatus
= Hydrosaurus pustulatus, the Sailfin Lizard (Water Dragon) from
the Phillipines and northwest Australia]
On the inner side of the tympanic processes there are two S-shaped three-sided bones
cemented to the base of the skull (S.) At the front they have a
compressed rounded head and at the back a swelling. Where their side surfaces meet, there
is a groove with an obtuse interior angle, beginning at the head and ending in the middle
of the depressed rear swelling. The front head is smooth, like a joint surface, and on the
bone to the right the head lies in the opening between the upper [??] and the tympanic
process of the pterygoid bone. The rough, notched rear end, on the other hand, indicates a
break, or a
Skull Structure of the Mosasaurus
(187) 15
connection with an apophysis. The length of these bones is 2 ¼ inches and their width
is one inch. They are either greatly elongated joint nodes of the sphenoid bone for the
reception of the pterygoid bone, or they are the columellae. However, they do not have the
necessary length to be the latter, and one would have to assume that they were extended by
an apophysis. Their considerable thickness would be in accord with the weight of the
occiput, for which they were to serve as a support.
The transverse bone is absent.
The boundary between the intermaxillary bone and the vomers (V.)
is not obvious. The latter are narrow, separated by a narrow groove, and they extend into
the area of the fourth molar, where they meet the narrow processes of the palatal bones (Plate VIII, Pa.) These do not
diverge farther from each other than the vomers; they are narrow and broken off at the
back, at the edge of the orbit. They resemble those of the Monitor, but have
a greater width, are closer together, and their outer edge joins with the maxilla to a
greater extent. The inner nasal cavities also resemble those of the Monitor,
but are narrower and shorter. In the Monitor they extend to the edge of the
orbit; in this specimen they end parallel to the exterior edge of the lachrymal bones. The
boundary between the vomers and the palatal bones is not visible.
The tympanic bone (Plate VIII, Tt.)
on the left side was shifted, and cemented to one branch of the mandible. Though it could
not be completely freed from the stony crust, one still sees that it is shell-shaped and
similar to that of the Maastricht animal. The curved process at the upper exterior corner
is not visible and is probably broken off.
The two rami of the mandible, which are broken off at the chin symphysis (Plate VII), agree in their outlines and in their structure
with those of Mosasaurus
Hofmanni.
16 (188)
A. Goldfuss
They could not be completely freed from the rock. There seem to be 10 teeth on each
ramus, and 4 teeth were thus probably lost with the broken-off piece, since the Maastricht
specimen has 14 teeth. The number of the nerve cavities can also not be determined. In
both species of this genus the mandible most resembles that of the Monitor;
it is low, elongated and has low lying joint surfaces. However, the end process of the
articular bone (d.) is shorter; the coronal process lies farther
back and is not formed solely by the coronal bone (c.) [=
coronoid], for the supra-angular bone (e.) extends all the way up to it and provides a
considerable breadth towards the back. The alveolar bone (a.) [=
dentary] is cut off at a slant at its posterior edge, in a straight line from front to
back. The most prominent characteristic of this jaw, also seen in the Maastricht specimen,
is the pronounced protrusion of the opercular bone (b.) [=
splenial] on the outer side, where it meets with the angular bone (e.)
in a vertical line. Their suture lies in a groove which follows the outer boundary between
the two bones to the front and the back. In Cuvier’s drawing (Tab. 18, Fig. 1) this
is covered by the fragments of the maxilla, which lie on top of it, but it is clear in the
plaster cast.
From the preceding observations it is clear that the genus Mosasaurus has
only the dental alveoli in common with the crocodile-like saurians, and only the bony
orbital ring in common with the fish-like saurians, thus being associated with the
surviving lizards, and it corresponds in its main form to the Monitor. Like
the Monitor, its head is elongated towards the front; it has large, long
nostrils, fused nasal bones, long pterygoid bones and a similar mandible. On the other
hand, like the Stellionas, Iguanas, Amaivas, Skinks, Chameleons and Geckos,
it has a simple medio-frontal bone.A widening of the
Skull Structure of the Mosasaurus
(189) 17
anterior frontal bone to a similar degree is found in the skinks and the Iguanas,
and in the chameleon this bone joins the posterior frontal bone, as in this specimen, to
form the orbital rim.*) The Istiurus has a similarly shaped, gnarled
lachrymal bone, and in Iguanas and Chameleons these bones are
also of a relatively small size. The zygomatic arch is just as narrow and weak in Anolis
and Podinema, and the Istiurus is characterized by a similar
parietal bone with a very short frontal surface. Palatal teeth are found in Lacertia,
Iguanas and some Skinks; there is a shell-shaped tympanic bone in Thorictis
crocodilinus, and a greatly abbreviated occiput in the Leguanas.
Given this manifold agreement of characteristics with other genera, there remain the
following features which are not found in other saurians, which are thus characteristic
for this genus.
1) The opercular bone of the mandible projects out farther on
its outer side than in any other genera.
2) The teeth are not wedged in, but have grown into the
alveoli through their sockets.
3) Above the row of nerve openings on the dental rim of the
maxilla there are two additional rows of similar openings.
4) The medio-frontal bone does not form a straight line
at its posterior edge, but rather narrows to a point, and in an opening at its tip it
receives a spoon-shaped process of the parietal bone, which is pierced by the frontal
opening.
5) The orbit is longer and lower than in the other Lacertia.
_________________________
*) According to a letter from von Meyer, he discovered the same structures in
his genus Metopias, from the Upper Triassic sandstone [Keuper] near
Stuttgart, whereas it is not the case in Mastodosaurus and Capitosaurus,
which belong to the same family.
18 (190)
A. Goldfuss
6) The narrow mastoid bone and the mastoid processes of the
parietal bone lie flat, so that their broad surface is facing upwards.
7) The pterygoid bones stand crowded together to the end of
their straight, almost parallel rows of teeth; their wider surfaces are directed towards
the outside and the inside. They bear the rows of teeth on their inner edge which
protrudes downward, and they have a third process parallel to the tooth edge.
8) The temporal bone makes a horizontal triangular widening at
its posterior end, and abuts the mastoid bone only with the posterior edge of this
triangle.
9) Lastly, the brain cavity is much shorter and narrower
than in any surviving lizards.
Of the spinal column of our skeleton 87 vertebrae are preserved, which together have a
length of 13 ½ feet, and in their form they closely agree with those of Mosasaurus Hofmanni,
which are well known through Cuvier’s description and illustration. *) The
transverse processes of the thoracic vertebrae have only one joint surface for the
reception of a simple rib head, and the caudal vertebrae differ from those of other
saurians in the lack of the articular processes and in the fusion of the haemal arches
with the body of the vertebrae, as is the case in fishes. The total number of the
vertebrae can not be determined from the existing remains, any more than Cuvier could do
so for Mosasaurus Hofmanni. To be sure, several series of vertebrae in their
natural order were associated with one another by their mode of burial; however, gaps are
apparent between them, and the last vertebrae of the tail are completely absent.
___________________________
*) Oss. foss. page 326, tab. 19, 20
Skull Structure of the Mosasaurus
(191) 19
Of the atlas only one bowed piece is present, cemented to the skull next to the
occipital foramen. Above it is an unidentifiable centra-like body, which might perhaps
belong to the axis. Then follow nine similarly shaped vertebrae which gradually increase
in size, of which the last seven were connected in a series. Their centra are longer than
wide, and wider than high. Their spinous processes slant backwards only a little; the same
is true for the horizontal transverse processes, which have less thickness in the first
two vertebrae, and which are compressed from top to bottom. In the following vertebrae
these processes gradually increase in thickness, and they are compressed from front to
back. The have four articular processes. In the first two vertebrae the anterior articular
processes are distinguished by a considerable length. In the middle of the lower side of
the centrum there is a short pointed spinous process, increasing in size up to the third
vertebra, then decreasing again, until at the 8th and 9th it is
reduced to a pair of low bumps. It is not perfectly round, but compressed at the base.
In the Maastricht remains Cuvier found a connected series of eleven such vertebrae with
a simple inferior spinous process (Tab. 19, Fig. 1.), but not the two with the doubled
bumps; thus the number of this division of vertebrae increases to 13. Of these 13
vertebrae only four belong to the neck, if, in accord with the structure of the Monitor,
four vertebrae without ribs follow the atlas and the axis. In our skeleton thus only the
third and fourth cervical vertebrae are complete, and the last two are missing, as well as
the first two thoraco-lumbar vertebrae.
The second series of vertebrae has all of the processes of the first, except for the
inferior spinous process. Cuvier found only five of this type (Tab.19, Fig.2),
20 (192)
A. Goldfuss
and was therefore forced to include a part of the caudal vertebrae among the thoracic
vertebrae and to assume that only a few of these latter had articular processes. Because
of this assumption the location of the pelvis became doubtful. This doubt is now resolved
by the present specimen. There are specifically 24 thoracic vertebrae with no spinous
processes or bumps. If one then includes the other nine vertebrae which do have inferior
spinous processes, that makes a total of at least 33 thoraco-lumbar vertebrae, while the Monitor
iavanicus has only 22. Lacertia ocellata has 25, but Scincus
variegatus has 37 of them. The heavy traverse processes of the whole series lie
horizontal and at right angles to the axis, and decrease gradually in size, whereas the
spinous processes achieve a greater width. All the thoraco-lumbar vertebrae seem to have
supported ribs, as in the Monitor, so that the lumbar vertebrae can not be
differentiated.
The following third series of our skeleton contains 11 vertebrae lacking articular
processes (Oss. foss. Tab.19, Fig. 3). Of these, only the first and largest is
separate; the others are connected in a regular sequence. Their centra gradually assume a
triangular shape; they are taller than wide and shorter than in the preceding series.
Their transverse processes are longer and more slender and point somewhat downwards and
backwards. Their spinous processes become so wide that they touch one another in the
anterior vertebrae and have only a narrow gap in the posterior vertebrae. The
anterior-most, found separated from the others, has much heavier processes, and it
indicates some distance between it and the others. Vertebrae with transverse processes
pointing upwards, such as Camper has depicted (Oss. foss. Tab. 20, Fig. 12, 13)
were not found. Therefore one would assume that these had their position in front of the
first isolated vertebra of our series, just as there is also evidently a gap between this
isolated vertebra and the
Skull Structure of the Mosasaurus
(193) 21
ones following. Cuvier indeed counts 38 vertebrae of this type, though he suspects
(loc. cit., p. 334) that this unusually large number might not come from one individual,
but from several. Accordingly, the number of the anterior caudal vertebrae remains
indeterminate.
The vertebrae of the subsequent fourth series, those of the central section of the
tail, still have small transverse processes which gradually become shorter and thinner,
but they differ from the preceding by the presence of the chevron bone, which is located
at the center of their lower side, and not on their posterior end, as in the Monitor.
In the remains of our specimen there is a series of 22 similarly shaped vertebrae, all of
which have a haemal arch. It cannot be determined whether this bone was articulated or, as
it seems, if it was fused in all of the vertebrae. This series is found in 8 pieces of
rock, whose surfaces match neither one another nor the preceding rocks, so at least 8
vertebrae are certainly missing. Accordingly, the medial tail had at least 30 vertebrae.
Their spinous processes are narrower than in the preceding series, and in the last few
they assume a vertical position. At the beginning the chevron bones point downwards at a
right angle, but gradually assume an acute angle closer to the centra of the vertebrae.
The vertebrae of the posterior portion of the tail no longer have transverse processes,
but they retain fused chevron bones (Oss. foss., Tab. 19, Fig. 6). The centrum
becomes shorter and more cylindrical. Their spinous processes grow narrower; in the
anterior vertebrae of this series they are still vertical, but in the more posterior they
tilt a bit forward. The chevron bone forms an acute angle with the axis and is elongated
so much that in each group of four they jut out over one another. Cuvier counted 44 of
these vertebrae in the
22 (194)
A. Goldfuss
Maastricht specimen. In our remains only 19 were found, in seven unconnected pieces of
rock, of which the two last are smaller by half than the preceding ones, which accordingly
indicates a large gap. Following Cuvier’s count, 25 vertebrae would be missing. The
vertebrae of the tip of the tail, in which the chevron bone is also absent, so that they
retain only a spinous process, are completely lost in our skeleton; even Cuvier found only
seven of them.
Thus by the examination of our skeleton, at least the number and shape of the cervical
vertebrae, of the thoracic and anterior caudal vertebrae, are established more precisely,
as well as the situation of the pelvis.
The following table of the various series of vertebrae, of which the vertebrae are
either present, or can be presumed to be missing, is supplemented by the measurement or
estimate of the length:
|
|
|
|
Number of |
Length |
Length |
|
|
|
|
vertebrae |
Feet |
Inches |
| The Skull |
|
|
Probable length of the missing tip |
--- |
--- |
4 |
|
|
|
Length of the existing part |
--- |
1 |
9 |
| |
|
|
|
|
|
|
| Vertebral Column |
|
|
Atlas and axis |
2 |
--- |
3? |
|
|
Cervical Vertebrae |
Existing medial vertebrae |
2 |
--- |
4 |
|
With inf. spinous process |
|
Missing posterior cervical vertebrae |
2 |
--- |
5 |
| A. With superior spinous
process |
|
Thoraco- lumbar vertebrae |
Missing front thoraco-lumbar vertebrae |
2 |
--- |
5 |
|
No inferior spinous process |
|
Existing anterior thoraco- lumbar vertebrae |
7 |
1 |
3 |
|
|
Other existing thoraco- lumbar vertebrae |
|
24 |
5 |
2 |
|
|
Missing sacral vertebrae |
|
2 |
--- |
5 |
| B. With the superior spinous process,
but without articular processes |
No inferior chevron processes |
1st series of caudal vertebrae |
Existing Missing
Existing |
1 19 ?
10 |
--- 3
1 |
2 ---
6 |
|
With chevron processes |
2nd series of caudal vertebrae |
Existing Missing |
22 8 |
2 1 |
10 1 |
| C. With the superior spinous process
but without articular or transverse processes |
With chevron processes |
3rd series of caudal vertebrae |
Existing Missing |
19 25 ? |
2 1 |
3 1 |
|
No chevron processes |
4th series of caudal vertebrae |
Missing |
12 ? |
--- |
6 |
|
|
|
Totals: |
157 |
23 |
9 |
Skull Structure of the Mosasaurus
(195) 23
Some small fragments of ribs, imbedded in the stone next to the vertebrae, are also
cylindrical in shape and striped, as in Mosasaurus Hofmanni.
Of the bones of the extremities also only scanty remains were found. Two pieces of
bone, fused together (Plate IX, Fig. 1) which form an
angular slanting joint surface, are parts of the scapula and the coracoid (Oss. foss.
Tab. 19, Fig. 15). On the inner surface depicted here one recognizes the coracoid (b.) by
the foramen above the joint surface. The two bones meet almost in a straight line, and
they are broken off at the common segment of the superior edge. On the outer side, which
could not be completely freed from the adhering stony mass, one can see the distinctly
elongated portion of the process which lies nearest to this opening. The coracoid thus
appears to have only one cut-out, as in the Lacertia and skinks,
and not two, as in the Monitor. The common joint in the bones also shows a
longer neck and is thicker than in the Monitor.
A fragment of bone (Plate IX, Fig. 3), which I consider to be the olecranon
of the right ulna, is characterized by pronounced exterior gnarls. A
third fragment (Plate IX, Fig. 2) is a
part of the outer edge of the left pubic bone (Oss. foss., tab. 19,
Fig. 10, 12) which extends from the anterior corner to a
point near the symphysis of the hip bone. The two gnarled raised areas of the outer edge
are peculiarly developed, and at the posterior corner of the inner fracture surface there
is a smooth area which belongs to the outer edge of the opening located there. In
addition, there are two more rounded joint ends of cylindrical hollow bones, which are
perhaps part of the radius. In comparison
24 (196)
A. Goldfuss
with the size of the skull and vertebrae, these remains of the extremities are rather
small.
If we attempt to draw conclusions about the living conditions of creatures of this
species from existing data concerning the discovery site and the structure of the
skeleton, we can be certain only that they were sea-dwellers and mighty carnivorous
predators. The first is shown by their place of burial, in Cretaceous formations which
were deposited in the sea. The latter is clear from the characteristics of their teeth.
Everything else can be inferred only with greater or lesser probability.
Since they lived in the sea, the toes of their feet were without doubt connected by
webs; however, the findings do not permit the assumption that they were fins, such as the
ichthyosaurs had. They swam primarily by means of the powerful strokes of their flattened,
oar-like tail. But they do not appear to have been fast swimmers, because for this a
shorter and more rigid back would have been necessary. The length and flexibility of their
back, as well as their tail, suggest rather, that they could also go on land and crawl on
sand bars and the shore like crocodiles. Their elastic and flexible bodies were able to
overcome obstacles in their path by means of serpentine windings, although their feet were
no longer or stronger than in the skinks. They sought and found food in the vicinity of
the shore, and only fish and mollusks were available. Their sharp teeth were suited to the
rending of large animal bodies, but not for tearing and crushing of bones and hard parts.
Since their teeth grew on sockets consisting mainly of loose bony substance, they lacked
the necessary rigidity for that, and the delicate sharp edge of their ridges,
Skull Structure of the Mosasaurus
(197) 25
which is always found undamaged, would have been damaged and worn down. Thus their
feeding was limited to cartilaginous fish. In their era rays and sharks were so numerous
in those waters that they had a superabundance of food, which they could seize not only at
the surface with their wide jaws, but also in the dark depths and in the mud of the
shore-line. The latter is clear from a seemingly insignificant condition, that is, the
triple rows of nerve-holes lying above one another at the snout, which occur only singly
in the related reptiles. These show that numerous nerve branches spread out in the nose
and make it suitable for rooting around. This arrangement also allows a conjecture as to
the nature of its body skin. The snout cannot have been armored with plates, since their
presence would contradict the purpose of the nerve holes. However, if the head, which
bears the largest armor plates in the Lacertia, was actually covered with
bare skin, then one may conclude that the rest of the body was also not armored, and that
the memory of the extinct ichthyosaur from the preceding epoch was renewed, so to speak,
in these creatures. This supposition has also not been disproved by any discovery of
traces of scales and plates, which could just as well have been preserved in these
skeletons as they were in other saurians and in the fish of this same era.
The smallness of the surface of the brain permits the conclusion that these animals had
a great tenacity of life and achieved a great age, but that they were neither
26 (198)
A. Goldfuss
excitable nor quick in their movements, and in this respect they lagged behind their
relatives of the present era.
They seem to have multiplied abundantly. This assumption is, to be sure, contradicted
by the rarity of discoveries of their skulls to date; but the stone quarries at Maastricht
have already produced so many teeth and vertebrae that most collections have been enriched
by them. In the fields near Aachen there are countless fragments of petrified teeth and
bones of these creatures lying around, and Prince Maximilian zu Wied reports *) that whole
skeletons are not seldom found in the Big Bend area of the Upper Missouri.
It would be extremely desirable to discover the skulls of the two gigantic
contemporaries of our species, the Iguanadon
and the Hylaeosaurus,
and to investigate whether such skulls would display similar anomalies in their structure.
But even lacking detailed knowledge of them, it is already clear that with the Cretaceous
Period there came a turning point in the development of saurians. To be sure, Nature was
still producing colossal forms, but at the same time was introducing the seeds which led
ultimately, after many changes of direction, to the more delicate forms which now
predominate.
__________________________
*) Journeys, Vol. II, p. 78.
____________________________
Skull Structure of the Mosasaurus
(199) 27
Explanation of the Illustrations
(Editor's Note: In some cases, the more contemporary name
for a bone is provided in brackets - [= dentary ] )
Plate
VI.
The skull from above, life size.
Plate
VII.
The same from the side, with the lower jaw.
Plate VIII.
The lower side of the skull.
The following abbreviations are identical in all three of these
plates.
Jm. Intermaxillary bone (Zwischenkiefer, Os intermaxillare)
[= Premaxilla ]
M. Superior Maxilla (Oberkiefer, Maxilla superior)
m. Fracture surface of the zygomatic arch process
(Jochbogenfortsatz)
n. Nasal bone (Nasenbein, Os nasale)
F. Medio-frontal bone (Hauptstirnbein, Os frontis medium)
[= Frontal ]
Fa. Anterior frontal bones (Vordere Stirnbeine, Ossa
frontalia anteriora) [= Prefrontal ]
fa. Opening in the anterior frontal bones above the orbit
Fp. Posterior frontal bones (Hintere Stirnbeine, Ossa
frontalia posteriora) [ = Postorbitalfrontal ]
L. Lachrymal bones (Tränenbeine, Ossa
lacrymalia) [= Anterior jugal ]
P. Parietal bone (Scheitelbein, Os parietale)
p. Parietal foramen (Scheitelbeinloch)
T. Temporal bones (Schläfenbeine, Ossa temporalia) [=
Squamosal ]
Tp. Petrose bones (Felsenbeine, Ossa petrosa) [= Prootic
]
Tt. Tympanic bones (Paukenbeine, Ossa tympanica) [= Quadrate
]
Tm. Mastoid bones (Zitzenbeine, Ossa mastoidea) [= Supratemporal
]
Z. Zygomatic bones (Jochbeine, Ossa zygomatica) [= Jugal
]
A. Plates of the orbit of the eye (Knochenring des Auges) [= Scleral
bones ]
Os. Superior occipital bones (Obere Hinterhauptsbeine, Ossa
occipitalia superiora) [= Orbitosplenoid ]
Ob. Basilar bone (Grundbein, Os basilare) [= Basioccipital
]
Ol. Lateral occipital bones ( Seitliche Hinterhauptsbeine, Ossa
occipitalia lateralia) [= paraoccipital process of the opistholic ]
Pt. Pterygoid bones (Flügelbeine, Ossa pterygoidea)
o. Transverse Process of Pterygoid
bones (Querbeinfortsatz)
l. Tympanic process of Pterygoid
bones (Paukenfortsatz)
28 (200)
A. Goldfuss,
Skull Structure of the Mosasaurus
m. Dental rim process (?)
(Zahnrandfortsatz)
o'. l'. m'. Fracture surfaces of these
processes (Bruchflächen dieser Fortsätze)
S. Either the columellae,
or the pterygoid processes of the sphenoid bone (Keilbein)
Pl. Palatine bones (Gaumenbeine, Ossa
palatina)
V. Vomers (Pflugscharbeine, Vomeres)
Lower Jaw (Unterkiefer)
a. Alveolar bone (Zahnstück, Os alveolare) [=
dentary ]
b. Opercular bone (Deckstück, Deckelstück, Os
operculare) [= splenial ]
c. Coronal bone (Kronenstück, Os coronale)
[= coronoid ]
d. Articular bone (Gelenkstück, Os articulare)
[= posterior angular ]
e. Inferior angular bone (Unteres Eckstück, Os
angulare) [= anterior angular ]
f. Superior angular bone (Oberes Eckstück, Os
supra-angulare) [= surangular ]
Tab. IX
1 a. Shoulder blade, and b. Left
coracoid (Schnabelbein)
2. Fragment of the left pubic bone
* Edge of the pubic opening
3. Olecranon of the right ulna
4. Jaw tooth of Mosasaurus Hofmanni (Kieferzahn)
5. Palatal tooth of the same (Gaumenzahn)
6. The enlarged ridge (Grat) of a tooth fragment
7. The pterygoid bones of Mosasaurus Hofmanni in
their natural location, reduced
______________________________
Erratum:
p. 180, line 4. Replace "Sm." with
"Jm."
[Note: This change was made in the translation.]
|




{kind=link}
{kind=link}
{kind=link}