
THE BIZARRE ‘WALRUS-TURTLES’ OF THE TRIASSIC
Text © 2000 Darren Naish
Copyright © 2000-2010 by Mike Everhart
Updated 03/09/2010
LEFT: "Placodus" Copyright © Dan Varner; used with permission of Dan Varner
|
PLACODONTS:
THE BIZARRE ‘WALRUS-TURTLES’ OF THE TRIASSIC Text © 2000 Darren Naish Copyright © 2000-2010 by Mike Everhart Updated 03/09/2010
LEFT: "Placodus" Copyright © Dan Varner; used with permission of Dan Varner |
CONTENTS:
Introducing the placodonts
Placodont morphology
What and how did placodonts eat?
Unarmoured vs. armoured placodonts
Cyamodontids - wide skulls, wide carapaces
Placochelyids, the ‘reptilian rays’
Bizarre and enigmatic, square-snouted Henodus
Placodonts that aren’t
Introducing the placodonts
Placodonts were peculiar, mollusc-eating marine reptiles known only from the Triassic of Europe and the Middle East. Looking something like a cross between a walrus and a turtle, they came in armoured and unarmoured varieties (see below) and were first discovered by Georg Münster in 1830. Seeking help from the great ichthyologist Louis Agassiz, Münster misidentified these teeth as those of pycnodont fishes1 and gave them the name Placodus. Richard Owen realised that Placodus represented a new group of reptiles, which he officially recognised in 1858 (Owen 1858). Variously allied over the years with synapsids, ichthyosaurs and araeoscelidans, placodonts have proved to be strongly modified members of the Sauropterygia, the position that was originally advocated for them when Owen created this group in 1860.
No known placodont was large: most are between 1 and 2 m long and the very largest forms may have reached 3 m. All were short-limbed robust animals of the shallow nearshore environment. Equipped with particularly dense bones and armour plates, live placodonts would have been negatively buoyant and, like other bottom-feeding marine vertebrates (e.g., some plesiosaurs, seacows), would have had no difficulty in remaining on the seafloor. As all other evidence indicates that placodonts were predators of benthic, shelled invertebrates (see below), this negative buoyancy would have been to their advantage.
 |
LEFT: Diversity in placodont body shape, exemplified by (top) Placodus, (left) Placochelys and (right) Henodus. |
Lacking flippers or a propulsive tail equipped for fast movement, placodonts would have been slow-moving and conceivably prone to predation from predatory marine reptiles and sharks. Being of small size (compared to giants like large ichthyosaurs and plesiosaurs) and lacking a specialised aquatic body shape, placodonts were probably amphibious, resting, breeding and (maybe) avoiding aquatic predators by taking to land. They were probably slow and clumsy on land: especially the turtle-like cyamodontoids, which tended to have very broad, flattened bodies and very short limbs. Placodonts were unique amongst sauropterygians in possessing armour - in the most specialised members of the armoured group, this was developed into an extravagant carapace over the body and the base of the tail. With a strongly dorsoventrally compressed body, a long whip-like tail, and a mollusc-eating lifestyle, the armoured placodont Psephoderma from the Norian and Rhaetian of Europe may have been a reptilian mimic of the rays that evolved sometime later in the early Jurassic. Like rays, perhaps these placodonts avoided predators by burying themselves in sand or gravel on the seafloor (Mazin and Pinna 1993).
Placodont armour was most probably for defensive purposes, though Frank Westphal (1976) has suggested that its main purpose was to improve hydrodynamic efficiency and that the presence of longitudinal keels represent the evolution of hydrodynamically advantageous features. Though this might be correct, the absence of any structures allowing fast movement in placodonts would suggest that adaptations beneficial to fast movement wouldn’t really be needed: moreover, features advantageous to self defence would be! Two juvenile Cyamodus specimens have been reported from the stomach contents of Lariosaurus (Tschanz 1989) and it has also been suggested that phytosaurs, large predatory fish like Saurichthys and pterosaurs may have preyed on small or juvenile placodonts (Mazin and Pinna 1993). However, as noted by Michael Benton (1990), why large, fully adult placodonts possessed such extensive armour is perhaps something of a mystery as none of the marine predators of the time seem capable of tackling them.
Both armoured and unarmoured placodonts first appear in the Scythian (Lower Triassic). The youngest known placodonts - members of the armoured family Placochelyidae - are from the Rhaetian, the last stage of the Upper Triassic. Placodonts clearly became extinct at or before the Triassic-Jurassic boundary, as did a number of other marine reptile groups (Bardet 1995), and they left no descendants.
Placodont morphology
Unarmoured placodonts had robust bodies that were rather square-shaped in cross section. Their necks were short and probably not very mobile and their tails were long, tapering, and only slightly laterally compressed. In all placodonts, the upper limb bones (humeri and femora) tend to be fairly slender while the lower parts of the limbs, hands and feet were short and appear weak. The pectoral and pelvic girdles of placodonts were composed of somewhat expanded bones located more underneath the body that to its side: the contacts with the vertebral column appear to have been quite weak. These features all suggest that placodonts were not good at supporting their weight when on land but that they had adequate musculature to use their limbs as steering or propulsive paddles. Digits on the hands and feet may have been encased by webbing. All placodonts had elongate transverse processes on their vertebrae and unarmoured forms have tall neural spines. Unarmoured placodonts (Placodus and Paraplacodus) also have accessory articulations on their vertebrae (a hyposphene-hypantrum system). There tends to be a well developed set of gastralia that curve upwards at their lateral tips to meet with the ribs. Like other primitive sauropterygians, placodonts have two-headed cervical ribs but dorsal ribs that only articulate with the transverse processes. Placodont ribs are thick and heavy.
 |
LEFT: Reconstructed section across middle of skeleton of Paraplacodus broili to show rectangular cross section. After Peyer and Kuhn-Schynder (1955). |
Armoured placodonts tended to have broad, flattened bodies (with low neural spines) and short, paddle-like limbs. This morphology was carried to an extreme in Henodus chelyops from the Upper Ladinian of SW Germany, a form in which the carapace was nearly twice as broad as long. In Psephoderma, an armoured placodont with a pointed rostrum, the tail was very long and whip-like and with a separate armour plate covering the base of the tail and the pelvis. Henodus had a shortened tail.
Placodonts had robust skulls that housed rounded, flattened teeth borne on the maxillae, palatines and dentaries. The bones of the palate and back of the skull are massive, robust and firmly wedged or fused together. Muscle attachment sites show that the adductor muscles, which appear tremendously well developed in all placodonts, ran largely horizontally, thus pulling the jaw backwards as it closed. The skull therefore appears suited for dissipating the strong forces generated during shell crushing.
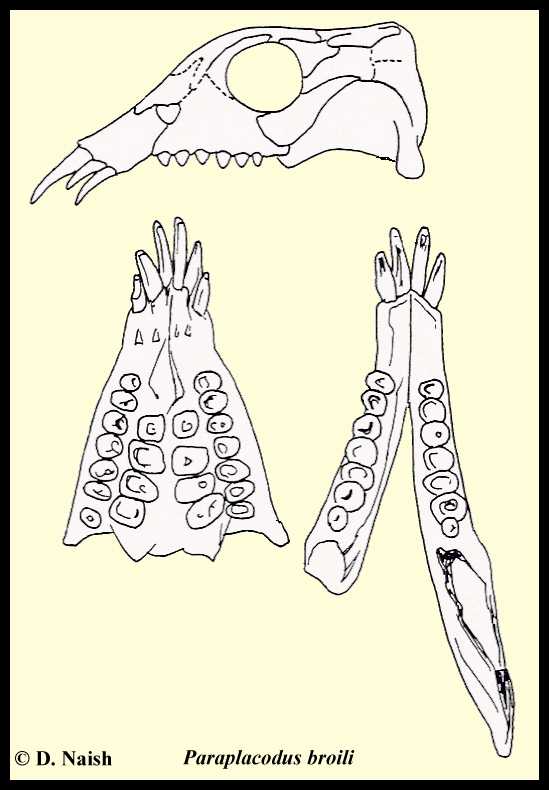 |
LEFT: Reconstruction of skull of Paraplacodus broili. After Reippel and Zanon (1997). (Below left): ventral view of Paraplacodus broili palate. After Peyer and Kuhn-Schynder (1955). (Below right) dorsal view of Paraplacodus broili lower jaw. After Peyer and Kuhn-Schynder (1955). |
Though advanced armoured placodonts lost their premaxillary teeth, nearly all other forms had protruding teeth at the front of the jaws which appear suited for plucking shelled invertebrates from the sea floor. In the unarmoured Placodus, the rostrum was broad and slightly expanded, supporting six robust spatulate teeth. This morphology appears powerful enough to have allowed Placodus to tear shelled prey from rocks and other hard surfaces. Premaxillary teeth were particularly elongate and prominent in Paraplacodus broilli, a poorly described species from the Grenzbitumen horizon (Anisian-Ladinian boundary) of Monte San Giorgio, Switzerland. The armoured placochelyids did not have premaxillary teeth, but instead had a pointed rostrum that does not appear robust enough to have pulled shelled prey off a hard substrate. They might therefore have extracted prey from mud or sand.
The pavement of crushing teeth at the back of the skull indicates that placodonts fed by crushing the shells of invertebrates and then feeding on the soft parts. So far as we know, no placodont ever lost all its teeth, and all of them appear to have depended upon this durophagous diet. The most highly modified form, Henodus, had an extremely unusual skull with an expanded, squared-off snout and only one pair of small teeth, located far to the rear on the palatine bones (though see below). In keeping with other features which indicate that it hunted by walking along the bottom of the lagoonal environment it inhabited, the square snout of Henodus may have been for grubbing around in the mud in search of invertebrates. Presumably its prey were softer than the organisms eaten by other placodonts.
What and how did placodonts eat?
With these remarkable anterior plucking and posterior crushing teeth, placodonts have usually been regarded as predators of bivalves. Presumably, they would crush their mollusc prey by positioning it in between their enormous palatal and lower jaw teeth, spit out the bits of shell, and then swallow the soft parts. By articulating the skulls and lower jaws of placodont fossils, it is possible to see that there was a main crushing area centred on the largest teeth of the palate, and a number of smaller ones situated around the smaller crushing teeth at the front of the palate.
Because of this probable lifestyle (the only one that is in keeping with the bizarre placodont morphology), placodonts have often been regarded as reptilian analogues of walruses (Odobenus). However, we now know that walruses do not crush their mollusc prey (it used to be thought that they did), but use a tremendous amount of suction to remove the extruding soft parts instead. Placodonts and walruses do not therefore seem to have fed in exactly the same manner.
Suggestions that placodonts may have fed on animals other than bivalves have been made from time to time. Emile Kuhn-Schnyder (1965) suggested that crustaceans may have been eaten. The extremely specialised and geographically restricted Henodus may have been a predator of Estheria, a relatively soft-bodied invertebrate that inhabited the same lagoonal habitat. A somewhat more problematic view is that placodonts were feeding on brachiopods. This latter idea was suggested by Freidrich von Huene who described broken masses of brachiopod shells as representing the remains of placodont meals (Huene 1933). It was also advocated by Bernard Peyer and Emile Kuhn-Schnyder (1955), simply on the basis of the abundance of brachiopods at placodont-bearing localities. I refer to this idea as ‘problematic’ because, as pointed out to me by Bristol University palaeobiologist Don Henderson after I mentioned this possibility during a talk, brachiopods appear to be remarkably unnutritious food items. Being immobile filter-feeding lophophorates with weak muscles and an expanded filter-feeding apparatus taking up most of the shell, brachiopods would hardly provide much of a meal for a hungry reptile. The subject therefore requires more thought and if you can help out here I’d be interested to hear from you!
Unarmoured vs. armoured placodonts
Placodonts can be divided into two major groups, the Placodontoidea Cope 1871 (not Peyer and Kuhn-Schnyder 1955) and Cyamodontoidea Nopsca 1923 (not Peyer and Kuhn-Schnyder 1955).
Placodontoids, sometimes called unarmoured placodonts, were less ‘modified’ than cyamodontoids, and still had a typically elongate reptilian body and a relatively tall and narrow skull. They also lacked an extensive covering of armour plates (though armour plates grew on the tops of their neural spines) and possessed a diastema separating their premaxillary teeth from their maxillary ones. Other salient features of their morphology were mentioned in the morphology section above. Only two placodontoid genera are presently recognised, Paraplacodus and Placodus.
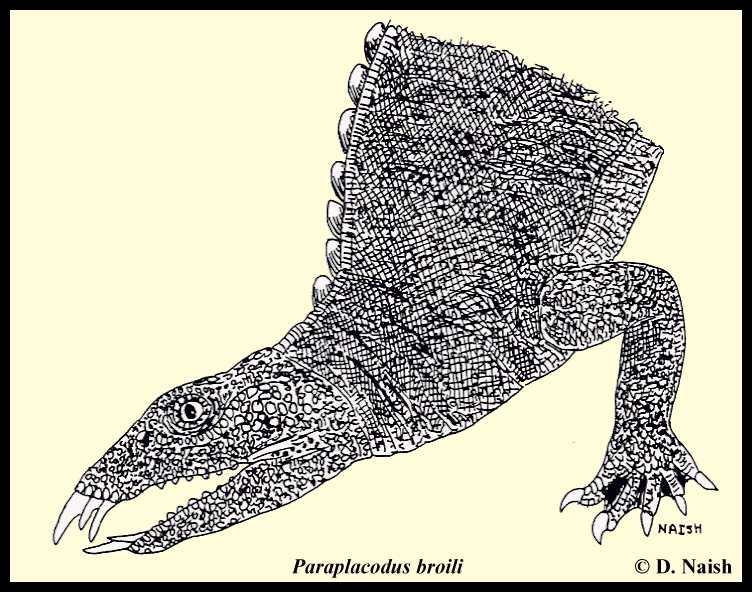 |
LEFT: Life restoration of Paraplacodus broili. |
 |
LEFT: Life restoration of Placodus gigas, based on a painting by Giovanni Caselli. |
Cyamodontoids, the armoured placodonts, had reduced or absent premaxillary teeth and had developed a turtle-like carapace composed of interlocking scutes. Armour plates also grew on the skull in cyamodontoids and the number of teeth both on the maxillae and the palatines was reduced. Cyamodontoids fall into three groups, traditionally regarded as families: the Cyamodontidae, the Placochelyidae and the Henodontidae.
Cyamodontids - wide skulls, wide carapaces
Cyamodontids are represented only by Cyamodus Meyer 1863, a genus for which six species from Anisian and Ladinian times have been named. Some of these species might be synonymous and Kuhn-Schnyder (1960) even suggested that Cyamodus be reserved for the type species, C. rostratus, while all other species be referred to a new genus. The geographically isolated C. hildegardis, recently described in detail by Giovanni Pinna (1992), may further represent a separate genus. The most recently recognised of these species, C. kuhnschnyderi Nosotti and Pinna 1993 (originally incorrectly written as C. kuhn-schnyderi (Nosotti and Pinna 1993a) - hyphens are not permitted in scientific names), may not be valid as its most distinctive feature [a sculptured lateral surface on the epipterygoid] is apparently an artifact caused by acid preparation (Rieppel in Rieppel and Zanon 1997).
Cyamodus has a remarkable skull that is anteroposteriorly short, with a reduced snout and face region, but incredibly wide, with enormous temporal fenestra. Emil Kuhn-Schnyder (1965) described the skull of Cyamodus rostratus as ‘heart shaped’. Depending on the species, there are either two or three rather small maxillary teeth and two or three palatine teeth (Mazin 1989), the most posterior of which is much larger than the more rostral one. Unlike the placochelyids, Cyamodus still possesses two small teeth in each premaxilla and these probably projected forward somewhat. Mazin (1989) described how juvenile specimens appear to have one extra tooth on the palate or maxilla compared to adults, and therefore individuals might reduce the number of teeth during their ontogeny. The cyamodont skull appears to have been very robust and well braced at the rear and Nosotti and Pinna (1993b) described features in this region which allow cyamodonts to be distinguished from placochelyids.
Cyamodus had a very broad, flattened body with a carapace consisting of hexagonal to subcircular osteoderms. The carapace was extensive, and covered much of the neck and virtually the entire span of the forelimbs. A separate caudal plate also covered the hips and base of the tail. The tail was stout and covered in osteoderms, as were the proximal portions of the limbs (Pinna 1992). Mazin and Pinna (1993) suggested that, because of its more strongly developed dentition, stronger limbs and deeper body, Cyamodus was less bottom-dependant and more mobile than the placochelyids, perhaps living in rougher waters in a more rocky environment.
An intriguing placodont that appears to be intermediate between Cyamodus and the placochelyids, Protenodontosaurus italicus, was described by Giovanni Pinna in 1990.
Placochelyids, the ‘reptilian rays’
Placochelyids comprise the two genera Placochelys and Psephoderma: two genera united by their notably triangular (in dorsal or ventral view) skulls that are not as transversely expanded posteriorly as that of Cyamodus, their pointed and edentulous rostra, and their even more extensive, turtle-like carapaces. Like Cyamodus, both genera have only two pairs of palatal teeth - a very large posterior pair, and a small rostral pair. Placochelys has three small, rounded teeth on each maxilla; Psephoderma has two, the posterior one being larger. As they were probably unable to pull prey from rocky substrates and had flattened bodies and very long, slim tails, placochelyids have been interpreted as reptilian ray mimics that may have hidden in the sea floor substrate (Mazin and Pinna 1993).
Psephosaurus is a problematic genus that was originally named for isolated scutes and carapace fragments from the Lower Keuper of Germany, but to which a great deal of material (particularly from Israel) was referred. The partial skull that is the type for P. mosis Brotzen, 1957 was used by Mazin (1989) as evidence that Psephosaurus is a placochelyid with a more reduced dentition than Placochelys or Psephoderma, but Rieppel and Zanon (1997) note both that referral of this skull to Psephosaurus is uncertain, and that it resembles Cyamodus. Westphal (1976) figures a great deal of Psephosaurus carapace material.
Bizarre and enigmatic, square-snouted Henodus
Henodontids, named for Henodus chelyops, are known only from Gipskeuper deposit of Tübingen-Lüstnau (Carnian, early Upper Triassic), southern Germany (Huene 1936). Believed to represent a semi-enclosed brackish or possibly freshwater lagoon, this environment shows that Henodus was the only placodont known to have inhabited a non-marine environment. Jean Michel Mazin and Giovanni Pinna (1993) suggest that H. chelyops represents a species that became isolated in this lagoonal basin: devoid of predators, and with a limited diversity of prey, it became supremely specialised for foraging in soft sediment on the lagoon floor.
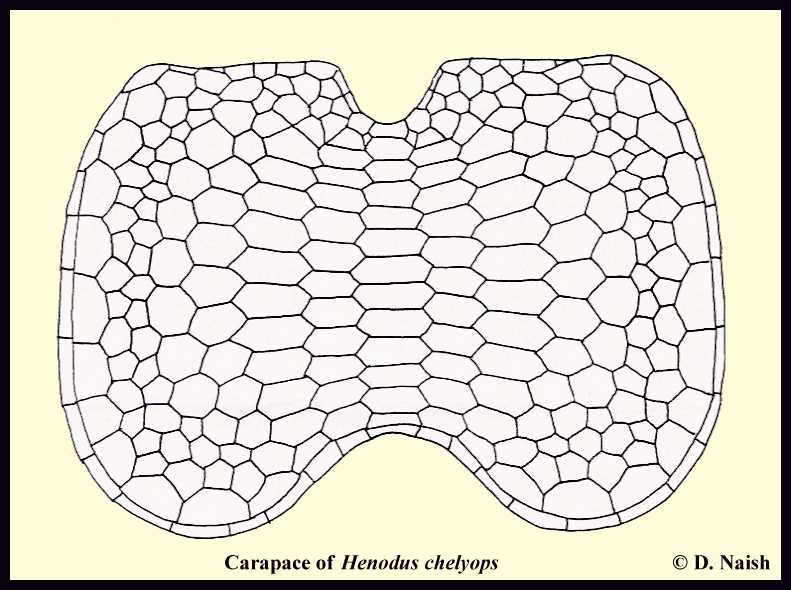 |
LEFT: Carapace of Henodus chelyops in dorsal view showing
arrangement of scutes. Click here for a color version. Click here for a computer generated Henodus from the National Geographic IMAX movie - Sea Monsters |
With its uniquely reduced dentition, square-snouted skull where the region in front of the eyes is tremendously shortened, and extensive carapace, Henodus appears to be most closely related to the placochelyids. Both share the presence of a plastron (a continuous armour shield on the underside of the body) and, supposedly, the absence of premaxillary teeth. However, a confusing and exciting discovery is that of a new Henodus specimen that has small and closely spaced premaxillary teeth (Hungerbühler et al. 1995): a discovery that suggests that Henodus may actually represent the primitive sister-group to all other cyamodontoids. If this is so, the late appearance of Henodus in the fossil record might be explained, suggest Oliver Rieppel and Robert Zanon (1997), by the fact that it was restricted to a peculiar environment that is not preserved in older rocks of the same region. At the time of writing (mid 1999), Oliver Rieppel is due to talk about placodont evolution at the 47th Symposium on Vertebrate Palaeontology and Comparative Anatomy. I look forward to hearing what he will say about this issue and will provide an update to this page after I have attended his talk.
Placodonts that aren’t
Two forms previously regarded as primitive, basal placodonts are no longer thought of as such. Helveticosaurus zollingeri from the Anisian of Switzerland was described by Peyer
and has traditionally been treated as the most primitive placodont of them all. Peyer and Kuhn-Schnyder (1955) regarded it as the sole member of a third placodont subgroup, the Helveticosauroidea, and Mazin (1989) classified it as the sister-group to all other placodonts. However, in 1989 Rieppel published his analysis of Helveticosaurus, and his conclusion that it was not a placodont: it is instead a bizarre kind of aquatic non-sauropterygian archosauromorph (Rieppel 1989).
Another supposed basal placodont, Negevodus ramonensis, was described by Mazin (1986) on the basis of a fragment of upper jaw. Unlike all other placodonts, Negevodus has a large, circular palatine fenestra (in other placodonts this is a narrow, slit aligned perpendicular to the skull’s long axis) and an external bone texture that, uniquely, is decorated with numerous small bumps. In his phylogenetic analysis of placodonts, Mazin (1989) regarded the position of Negevodus as problematic because it combined a reduced palatine dentition (a cyamodontoid condition) with a full developed maxillary dentition (a placodontoid condition). Mazin therefore suggested three possibilities: either Negevodus was a placodontoid that had convergently evolved a cyamodontoid-like palatal dentition, it was a peculiar cyamodontoid that had reacquired the primitive placodontoid-type maxillary arrangement, OR (in view of its totally unique huge palatine fenestra) it represented a new type of very primitive placodont that had convergently evolved cyamodontoid-like palatal teeth. The numerous un-placodont like features were always cause for concern, however, and these led Robert Zanon, in 1991, to reinterpret the specimen as a temnospondyl amphibian (Zanon 1991).
1.
Pycnodonts are deep-bodied fossil neopterygian fishes from the Mesozoic and Eocene that, like placodonts, had large and rounded teeth on their palates and lower jaws.References
{kind=link}
{kind=link}