 |
Plesiosaur Stomach Contents and
Gastroliths from the Pierre Shale (Middle Campanian) of Kansas
David J. Cicimurri
Bob Campbell Geology Museum, Clemson University, Clemson, S.C.
29634-5210
and
Michael J. Everhart
Sternberg Museum of Natural History, Fort Hays State University,
Hays, KS, 67601
Copyright © 2001-2009 by Mike Everhart
Webpage updated 09/18/2009 |
AUTHORS NOTE: Do not cite or quote
from this webpage. This is a early draft of the published manuscript, with additional or
updated figures and references. Please refer to: Cicimurri, D. J. and M. J. Everhart,
2001. An elasmosaur with stomach contents and gastroliths from the Pierre Shale (late
Cretaceous) of Kansas. Kansas Acad. Sci. Trans 104(3-4):129-143. "Feeding Styxosaurus"
(above) is copyright © 2001 by Russell Hawley.
Click here for additional pictures of the
NJSM 15435 gastroliths and stomach contents.

_________________________________
Abstract
A nearly complete skeleton of an elasmosaurid plesiosaur (NJSM 15435) from the
Sharon Springs Member (Middle Campanian) of the Pierre Shale, Logan County, Kansas, is
intimately associated with fragmentary fish remains and numerous gastroliths. The fish
bones and gastroliths were located just behind the pectoral girdle in the abdominal
region. Identifiable prey includes Enchodus and other small clupeomorph fishes. An
isolated tooth of the anacoracid shark Squalicorax cf. S. pristodontus
was also recovered in this area. Ninety-five gastroliths (6.8 kg) were present, with the
largest stone measuring 15.1 x 8.5 x 5.7 cm (5.0 x 3.3 x 2.2 in.) and weighing 1.06 kg
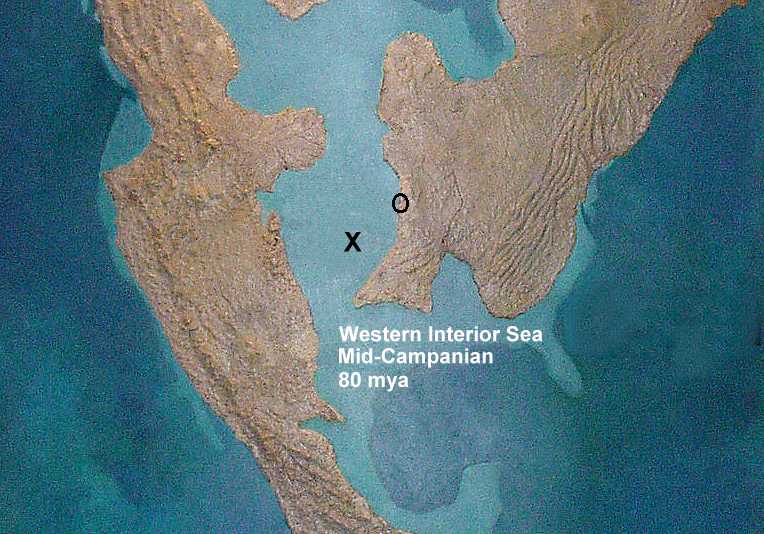
(2.3 lb.). Many of the gastroliths are composed of pink or gray Sioux Quartzite, which suggests that the source of these stones (O) was over
600 km (475 mi.) to the northeast of where the elasmosaur remains (X) were discovered.
The association of fragmentary fish remains and gastroliths within the abdomen of NJSM
15435 supports the contention that the stones aided in the breakdown of food in
plesiosaurs.
________________________________
Introduction
Determination of the diet of extinct animals from the fossil record is usually
indirect, especially within a marine ecosystem. One can infer dietary preferences by
comparing tooth morphology to that of extant animals (Massare, 1987). Shed teeth and cut
marks on bone can indicate that certain species of sharks fed upon fish and marine reptile
carcasses (Schwimmer et al.; 1997; Shimada, 1997; Everhart, 1999). Some food items
may be the result of opportunistic scavenging, and may not necessarily represent the
typical prey for those particular sharks (Schwimmer, 1997). Coprolites offer an accurate
glimpse into the diet of fossil vertebrates, but it is often difficult to identify the
predator taxon (Martin and Bjork, 1987). For that reason, gastric residues or stomach
contents associated with fossil skeletons are particularly valuable because both the
predator and prey taxa can often be identified.
Occurrences of fossil vertebrates with preserved stomach contents are rare.
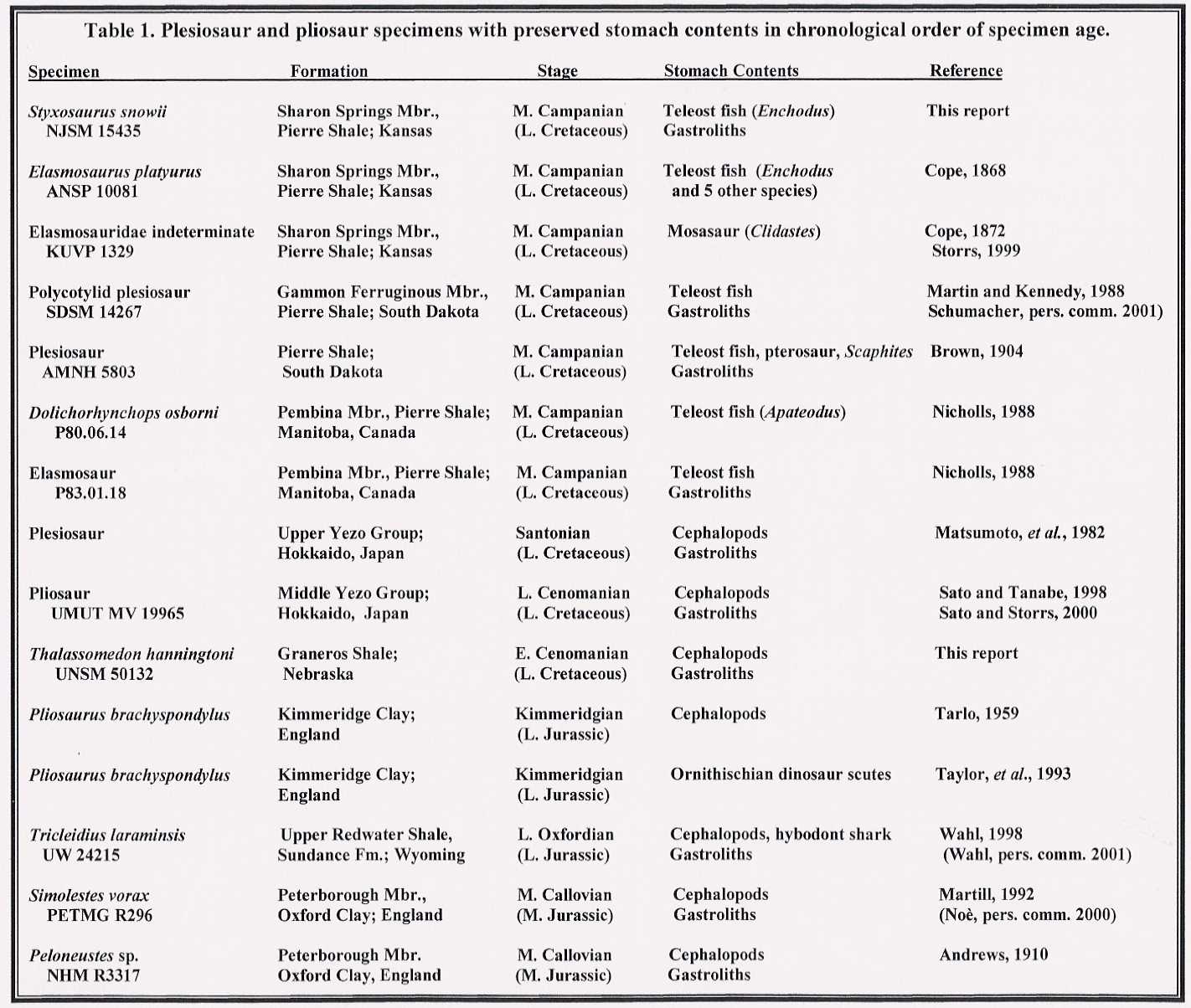
While gastroliths are often associated with plesiosaur skeletons (Williston, 1904, 1914),
only six sets of plesiosaur remains containing gastric residues have previously been
reported from the late Cretaceous Pierre Shale (Table 1).
The vertebrae, scales, and teeth of six species of fish, including Enchodus, were
associated with the type specimen of Elasmosaurus platyurus (ANSP 10081) discovered
by Captain Theophilus Turner, an Army physician at Fort Wallace, in the Pierre Shale of
Logan County, Kansas in 1867 (Cope, 1868). Cope (1872) reported the remains of a juvenile
mosasaur (Clidastes) beneath the vertebral column of a much larger elasmosaur (the
type of Plesiosaurus gulo) from the Pierre Shale near McAllaster, Logan County,
Kansas (see also Storrs, 1999, for the identification and stratigraphic occurrence of this
specimen). Brown (1904) described plesiosaur remains (AMNH 5803) from the Ft. Pierre
Formation, at the "head of Hat Creek Basin, 18 mi. southwest of Edgemont, South
Dakota" that contained fish vertebrae, broken pterodactyl bones, and Scaphites
as stomach contents. Martin and Kennedy (1988) documented the remains of a plesiosaur from
the Pierre Shale of South Dakota (SDSM 14267) with stomach contents (teleost fish
vertebrae and scales) and gastroliths. A specimen of Trinacromerum bonneri (now Dolichorhynchops
osborni) containing teeth of the enchodontid Apateodus as gastric residue, and
an elasmosaur associated with gastroliths and macerated fish remains were reported by
Nicholls (1988) from the Pembina Member of the Pierre Shale in Manitoba, Canada.
In a general way, records of pre-Campanian plesiosaur food items (Table 1) tend to include mostly cephalopods (squid,
belemnite and ammonite) and few or no fish. Cephalopod beaks associated with gastroliths
were preserved as stomach contents in a pliosaur specimen from the late Cenomanian of
Japan (Sato and Tanabe, 1998; Sato and Storrs, 2000). An elasmosaur skeleton from
the early Cenomanian of Nebraska (Thalassomedon haningtoni Welles 1943, UNSM 50132)
at the University of Nebraska State Museum includes fragments of ammonite shells mixed
with gastroliths in the abdominal region. Tarlo (1959) documented cephalopod hooklets
associated with the remains of Pliosaurus brachyspondylus from the late Jurassic
Kimmeridge Clay of England. Taylor, et al. (1993) described the dermal scutes of an
ornithischian dinosaur associated with another specimen of the pliosaur Pliosaurus
brachyspondylus. Wahl (1998) reported the remains of a hybodont shark (teeth, dorsal
spines and skin denticles) and coleoid cephalopod hooklets from a late Jurassic plesiosaur
skeleton from Wyoming.
Fossilized remains of marine reptiles other than plesiosaurs have also included
stomach contents. Martin and Bjork (1987) described the remains of a shark (Cretalamna(?)
teeth), a teleost fish (Bananogmius), a marine bird (Hesperornis) and
another mosasaur (Clidastes) in the preserved stomach contents of a large Tylosaurus
proriger from the Pierre Shale of South Dakota. Camp (1942) reported teleost fish
remains associated with a California mosasaur (Plotosaurus tuckeri Camp 1942, CIT
2750). Everhart (in preparation) observed partially digested fish vertebrae (Cimolichthys
nepaholica) and scales in the abdominal region of a 10 m (33 ft.) Tylosaurus
proriger skeleton (FFHM 1997-10) from the Smoky Hill Chalk Formation, Gove County,
Kansas. Pollard (1968) recovered cephalopod hooklets mixed with small quartz grains from
ichthyosaur remains discovered in the Lower Lias (early Jurassic) of Lyme Regis, England.
The rarity of such specimens may result from several factors, including: the
decay and disarticulation of the carcass prior to burial (Schäfer, 1972; Nicholls, 1988;
Schwimmer, 1997; Everhart, 2000); the scavenging of the carcasses by sharks which results
in the removal of the visceral elements (Shimada, 1997); or the death of the animal long
after the last ingestion of food.
Abbreviations
The abbreviations for institutions referred to in the text are as follows: AMNH
– American Museum of Natural History, New York; ANSP – Academy of Natural
Sciences of Philadelphia; CIT - California Institute of Technology, Pasadena, CA; FFHM
– Fick Fossil and History Museum, Oakley, KS; FMNH – Field Museum of Natural
History, Chicago, IL; KUVP – University of Kansas Vertebrate Paleontology Collection,
Lawrence, KS; NJSM – New Jersey State Museum, Trenton, NJ; NZGS – New Zealand
Geological Survey, Lower Hutt, NZ; SDSM South Dakota School of Mines and Technology, Rapid
City, SD; SMUSMP – Southern Methodist University, Shuler Museum of Paleontology,
Dallas, TX; TMM – Texas Memorial Museum, University of Texas at Austin; UMUT –
University Museum, University of Tokyo, Japan; UNSM – University of Nebraska State
Museum, Lincoln, NE.
Geographic and Stratigraphic Occurrence
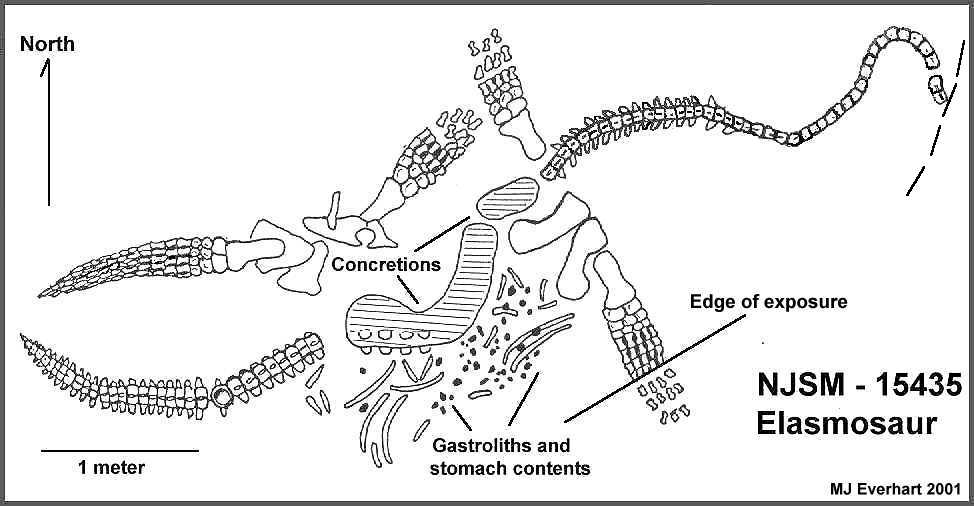
NJSM 15435 was discovered in 1991 by Pete Bussen, a local rancher, from
elements of the right front limb eroding from the edge of a gully in an area locally
referred to as "Coal Oil Canyon" in western Logan County, Kansas. The plesiosaur
was lying along a northeast to southwest axis (Figure 1).
Dr. David Parris of the New Jersey State Museum was offered the opportunity to dig the
remains as the result of an earlier contact with Mr. Bussen. Staff
and volunteers from the NJSM recovered the specimen on three successive digs in 1991-1992. Stratigraphically, the specimen was located below the
mid-point of the Sharon Springs Member of the Pierre Shale (Baculites maclearni [mclearni] biozone). The type section of this member is
located about 22 km northeast of the excavation, near McAllaster Buttes, and is named for
the nearby town of Sharon Springs, Kansas (Elias, 1931). The Sharon Springs Member was
deposited in a shallow inland sea that covered much of the central United States and
Canada during middle Campanian time (Parrish and Gaultier, 1993). This lithostratigraphic
unit is composed of soft, fissile, bituminous shale and contains numerous bentonites and
septarian concretions (Gill et al., 1972). Extensive field work in the Sharon
Springs Member in South Dakota by the SDSMT has documented a diverse vertebrate assemblage
consisting of fish, marine turtles, plesiosaurs, mosasaurs, pterosaurs, and birds (Martin
and Bjork, 1987). Although such rigorous surveys have not yet been conducted in the Sharon
Springs Member in Kansas, the same groups of vertebrates are also well represented
(Carpenter, 1990).
* Postscript: the proper spelling of the species
name, contrary to Cobban (1993), is mclearni; from B. mclearni Landes
1940
Taphonomy and Preservation of the Specimen
NJSM 15435 (Figure 1) represents the nearly
complete skeleton of a large elasmosaurid plesiosaur that was tentatively identified in
the field as "Alzadasaurus snowii." on the basis of the characteristic
shape of the pelvic girdle (D. Parris, pers. comm., 1991). Since that time, however,
Carpenter (1999) has proposed that Alzadasaurus is a junior synonym of Styxosaurus.
Field measurements of the complete and fully articulated
left rear limb (1.7 m) and caudal vertebrae (2.0 m) indicate that the plesiosaur was
about 10 m (33 ft.) in length and would have weighed over 2,800 kg (Everhart, 2000). The
specimen includes nearly complete pectoral and pelvic girdles, most of the vertebral
column, ribs and gastralia, and almost all of the limb elements. Unfortunately, surface
erosion prior to the discovery of the remains had removed the area of shale that may have
contained the anterior-most portion of the neck and the skull. Gastroliths, teleost fish bones, teeth and scales were
located in an area of discolored shale between the pectoral and pelvic girdles on the
right side of the skeleton. The position of the gastroliths and the ribs, and the
location of the pectoral girdle in relation to the right front limb, indicate that NJSM
15435 came to rest on its chest. The right rear limb and pelvic girdle elements were
displaced anomalously to the left side of the vertebral column before fossilization. While
the distal ends of the right rear limb and left front limb are missing (possibly lost to
scavengers), the right front limb appeared to have been essentially complete prior to
eroding out of the shale. Although the ribs and gastralia were somewhat scattered within
the abdominal region of the elasmosaur, the relative completeness of the skeleton,
especially the left rear limb, suggests that the animal settled on the bottom shortly
after death and that significant scavenging did not occur. Scavenging may have been
inhibited if, as suggested by Parrish and Gaultier (1993), oxygen levels were reduced near
the sea floor. Articulated limbs, girdle elements, and vertebrae suggest that currents
were absent or insufficient to disturb the remains. The
scattering of ribs, gastralia and gastroliths within a relatively small area indicates
that the gut may have bloated and burst while the carcass was lying on the bottom. Most of
the dorsal vertebrae were enclosed in a series of limestone concretions. The concretions
may have formed where they did as the result of carbonate ions being drawn to a locally
restricted reducing environment produced by the decay of the gut (E. Manning, pers. comm.
2001).
Conditions were especially favorable within the Sharon Springs Member of the
Pierre Shale in Kansas for the secondary formation of selenite. As a result of anoxic
bottom conditions during and after burial, pyrite crystals were likely present throughout
both the bones and the surrounding clay. They provided the sulfur which, when combined
with the calcium from dissolved aragonitic bivalve shells, produced the calcium sulfate
crystals (E. Manning, pers. comm., 2001). These crystals can grow within bones and often
result in poorly preserved, highly fragmented and fragile fossils. There was substantial
degradation of the smaller bones of NJSM 15435 by the growth of selenite crystals. In this
instance, even the large, dense propodials were affected.
Stomach Contents
A fish bone hash was concentrated in a one by two-meter area just behind the
pectoral girdle on the right side of NJSM 15435. Gastroliths
and gastralia were scattered randomly throughout the bone hash within this area. The
gray shale in this region was stained red-orange, a diagenetic artifact possibly related
to the decomposition of the carcass. Matrix containing the stomach contents was collected
in two plaster jackets in 1992. Disarticulated fish
vertebrae, fragments of fish bone and scales, as well as numerous small (1-2 mm) flecks of
fish bone were dispersed throughout the discolored matrix (Figure 2). Small pieces of pulverized fish bone or scale
adhered to some the gastroliths. Many of the ribs and gastralia were located in a layer
under the gastroliths. There were no occurrences of gastroliths beneath this lower layer
of bones. Nicholls (1988) reported macerated fish bone associated with several plesiosaur
skeletons from the Pembina Member of the Pierre Shale, with fragments ranging in size from
1 mm to 3 cm. According to Nicholls (ibid.), the surface of these bone fragments showed no
evidence of chemical corrosion by stomach acids.
Fragments of fish bone were the most common elements recovered from the stomach
contents of NJSM 15435 during preparation. Materials that were considered to be diagnostic
and were sent to J. D. Stewart for identification. Cranial fragments of small clupeomorph
fishes were present and several teeth, vertebrae, and scales of the common Cretaceous
teleost Enchodus sp. Agassiz, 1835 were also identified (J. Stewart, pers. comm.,
1996). A nearly complete tooth of the shark Squalicorax cf. S. pristodontus
was also recovered from this area, as was an isolated, but unidentified, elasmobranch
vertebra. Storrs (1981) reported shark teeth associated with the thoracic ribs, gastralia
and gastroliths of a Thalassomedon sp. (TMM 42245-1) plesiosaur and concluded that
it was unlikely that the shark teeth were part of the stomach contents. As only one shark
tooth was recovered with NJSM 15435, it is regarded here only as possible evidence of
scavenging.
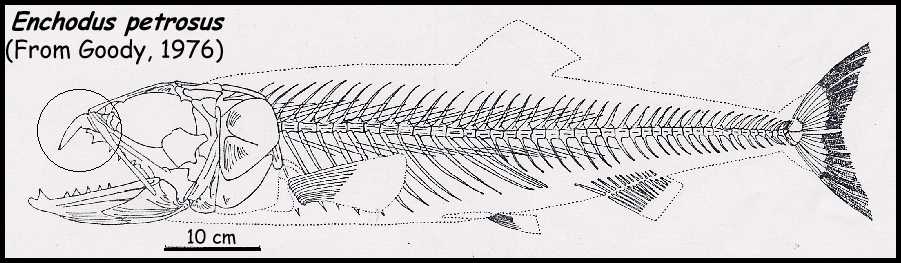
Enchodus
remains are common throughout the Smoky Hill Chalk and Pierre Shale. Based on measurements
of a complete Enchodus skeleton (Pete Bussen, private collection, pers. comm.,
2000) preserved as stomach contents inside a larger fish (Cimolichthys nepaholica)
from the Pierre Shale, a 75 cm (30 in) Enchodus has vertebrae with an average
length of 15 mm and diameter of 10 mm. Most of the fish
vertebrae associated with NJSM 15435 have a length of about 8 mm and a diameter of about 6
mm, suggesting a prey length of 35 to 45 cm (14 to 18 in).
Gastroliths
Lithophagy (stone swallowing) is well documented in many groups of animals (see
review in Whittle and Everhart, 2000) including plesiosaurs and dinosaurs. Seeley (1877)
reported "about a peck (1/3 cu. ft.) of ovate and rounded pebbles" in the
abdominal region of a Mauisaurus gardneri from England. Williston (1904) noted
"at least 30 instances" of gastroliths associated with the remains of
plesiosaurs from Europe and North America. "Small polished and rounded pebbles"
were reported by Ji, et al. (1998) in the crop of a feathered dinosaur,
Caudipteryx zoui. from China. Taylor (1999) concluded that Caudipteryx was
insectivorous and used gastroliths to grind up the hard exoskeletons of its prey.
Kobayashi, et al. (1999) documented masses of small siliceous grains in the crops
of 12 associated ornithomimid dinosaur skeletons. While the function of gastroliths in
plesiosaurs is still uncertain, it is probable that the grains served a similar function
in the digestive process of these dinosaurs as the grit in the crops of some modern birds.
 |
Ninety-five gastroliths were discovered with NJSM 15435. This is
far fewer than the 253 gastroliths reported by Welles and Bump (1949) in association with
an Alzadasaurus pembertoni (SDSM 451, now Styxosaurus snowii, Carpenter,
1999) from the Sharon Springs Member in South Dakota. There were 197 gastroliths recovered
from an elasmosaur in the Bearpaw Shale of Montana (Darby and Ojakangas, 1980), and
206 were documented by Riggs (1939) from a specimen of Hydralmosaurus serpentinus (FMNH
12009). Shuler (1950) reported that more than 70 dark flint stones were discovered within
the ribs of the nearly complete, type specimen of Libonectes morgani (SMUSMP
69120) from the early Turonian Britton Fm., Eagle Ford Group of Texas. In his description
of a Thalassomedon sp. skeleton (TMM 42245-1) that included 45 gastroliths, Storrs
(1981) concluded from the condition of the remains that the original number of stones had
been reduced through pre- and post-depositional erosion of the skeleton. Nicholls (1988)
reported "more than fifty" gastroliths associated with an elasmosaur skeleton
from the Pembina Shale, Manitoba, Canada. Forty-seven
unusually large gastroliths were associated with the incomplete remains of another
elasmosaur (KUVP 129744) discovered within 20 km of and at the same stratigraphic
level as NJSM 15435 (Everhart, 2000). |
Many of the NJSM 15435 gastroliths are pink quartzite. Other materials include
gray quartzite, quartz, chert, and sandstone. Williston (1893) reported red quartzite
gastroliths from a large Kansas plesiosaur from Ellsworth County, and concluded that they
were lithologically identical to Sioux Quartzite outcroppings in northwestern Iowa. Welles
and Bump (1949) described gastroliths associated with the type specimen of Alzadasaurus pembertoni (SDSM 451), now Styxosaurus
snowii, as being composed of pink and gray quartzite, siltstone, sandstone, and flint.
The lithology of the gastroliths from NJSM 15435 appears to be similar to that of SDSM
451, and it is possible that these plesiosaurs acquired their stones from the same general
geographic areas. Based on the presence of pink and gray quartzite, the source of some of
the stones may have been from islands or other localities along the eastern shore of the
Western Interior Sea where Precambrian quartzites were exposed during the time of
deposition of the Sharon Springs Member (Bretz, 1987). The gastroliths reported by Darby
and Ojakangas (1980) from the middle Campanian Bearpaw Shale were composed of gray
quartzite, chert, granite, basalt and other volcanics, and silicified shale, which
suggests different source areas than for NJSM 15435 and SDSM 451. Riggs (1939) reported
that all of the gastroliths in the specimen he studied from the Fort Benton Shale of
Montana were composed of granite. Most of the gastroliths in TMM 42245-1 (Storrs, 1981)
from the early Turonian of Texas were composed of a dark chert, with a few others
identified as siltstone and quartzite.
The NJSM 15435 gastroliths have a combined weight of approximately 6.8 kg (15
lb.). The largest gastrolith weighs 1,060 g (2.5 lb.), whereas the smallest weighs only
0.4 g. See Everhart (2000) for a comparison of the sizes and
weights of the NJSM 15435 gastroliths with those of other Late Cretaceous plesiosaurs.
Long/intermediate/short axis measurements were taken with dial calipers. The
largest gastrolith from NJSM 15435 measures 151 x 85.4 x 57.4 mm, and the smallest 8.6 x
6.6 x 5.9 mm. The average size of the gastroliths was about 30 x 25 x 15 mm. The
measurements of the intermediate and short axes of the largest gastrolith (85.4 x 57.4 mm)
of NJSM 15435 may provide an idea of the cross-sectional area of the largest prey that
could have been ingested by this elasmosaur. Shuler (1950) noted that a similar sized
specimen of Libonectes morgani would have only been able to swallow a fish with a
cross section no larger than 13 cm (5 in) because on the narrow width and inflexible
articulation (fused symphysis) of the lower jaws. According to Massare (1987), the maximum
skull width of many plesiosaurids was < 10 cm (4 in.). The inside of the reconstructed
lower jaw of Thalassomedon haningtoni (DMNH 1588) at the Denver Museum measures
about 15 cm (6 inches) wide where it articulates with the quadrates. Despite the huge size
of their bodies, the relatively small skull size and slender, seizing teeth suggests that
food items for elasmosaurs were limited to small diameter fish and soft-bodied
invertebrates.
Although the shape of the gastroliths of NJSM 15435 varies, they can be
categorized into three general types: disks (near equal long and intermediate-axis length,
shorter short-axis length), spheres (near equal long/intermediate/short-axis lengths), and
cylinders (greater long-axis length, near equal intermediate and short axis length). The
majority of the stones are well rounded, with smooth surfaces, and even those with flat
sides have rounded corners. They also have a thin exterior coating of iron oxide-stained
clay that tends to obscure the composition of the stone and the degree of polish. The
gastroliths composed of quartzite have a dull pitted texture, whereas those of chert are
highly polished. Those identified as quartz are dull and smooth. These observations appear
to correlate well with those of other authors (Welles and Bump, 1949; Darby and Ojakangas,
1980; Storrs, 1981; Wiffen and Moisley, 1986). The cause of the polishing has usually been
attributed to grinding against each other in the stomach or crop of the animal (Shuler,
1950; Gillette, 1990).
The function of gastroliths in plesiosaurs has been and continues to be a
controversial topic. Benjamin F. Mudge (1878), the first State
Geologist of Kansas, noted that "in the Plesiosauri, we found another interesting
feature, showing an aid to digestion similar to many living reptiles and some birds. This
consisted of well worn siliceous pebbles, from one-fourth to one-half inch in diameter.
They were the more curious, as we never found such pebbles in the chalk or shales of the
Niobrara." Because plesiosaurs probably swallowed their prey whole, these stones may
have been utilized in the mechanical breakdown of food (Brown, 1904; Andrews, 1910;
Williston, 1914; Shuler, 1950; Martin and Kennedy, 1988). In contrast, the fossilized
remains of mosasaurs regurgitated by large sharks (Everhart, 1999), predatory fish
preserved with intact prey such as the famous "fish in a fish" Xiphactinus specimen (FHSM VP-333) with a Gillicus (FHSM
VP-334) inside at the Sternberg Museum of Natural History,
and a mosasaur (Martin and Bjork, 1987)
with preserved stomach contents from the Western Interior Sea all suggest that prey,
or parts of prey, swallowed whole tends to remain more or less articulated during the
early stages of digestion in animals without gastroliths.
Cott (1961) concluded that extant crocodiles use stones for buoyancy
regulation, and Denton et al. (1997) proposed that this was the case in the extinct
Cretaceous/ Paleocene marine crocodile Hyposaurus. Many
authors have suggested that plesiosaurs used gastroliths to adjust buoyancy
(Williston, 1893; Darby and Ojakangas, 1980; Taylor, 1981; Storrs, 1981). Currie (1981)
demonstrated that the weight of the gastroliths in the lower abdomen of the aquatic
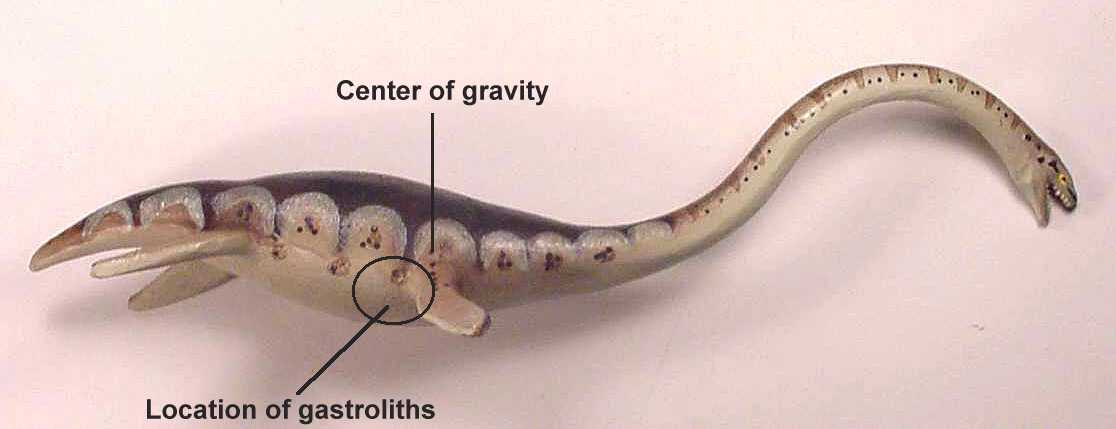
eosuchian, Hovasaurus boulie, would have lowered the center of gravity, increased
the specific gravity of the animal, and aided in maintaining an upright orientation for
swimming. Carroll (1985) proposed that the use of gastroliths and heavy gastralia for
buoyancy control in plesiosaurs was one of several adaptations made by diapsid reptiles
returning to life in the ocean. Reiss and Frey (1991) reviewed the evolution of underwater
flight in a variety of animals and concluded that ballasting would not have been required
for underwater flight in plesiosaurs. Storrs (1993) suggested that plesiosaur gastroliths
functioned in a dual role of helping to maintain a neutrally buoyant position in the water
column while also serving as a grinding aid to digestion. Sander et al. (1997)
suggested that a mass of sand grains discovered in the abdomen of a pistosaurid
sauropterygian from Triassic marine sediments in Nevada was used for buoyancy control.
Taylor (1993, 1994) discussed the physics of buoyancy in marine animals, and
proposed that a relatively small mass of these stones would be sufficient to influence the
position of the animal in the water column. Taylor (1993) also noted that gastroliths can
be quickly swallowed or vomited by some modern marine mammals to adjust buoyancy as
needed. It is doubtful, however, that this would have been a useful strategy for an
elasmosaur with a 5-6 m (16-20 ft.) long neck, especially one that was living and feeding
hundreds of kilometers from the nearest sources of such stones. Lingham-Soliar (2000)
suggested that gastroliths may have been used more to adjust the longitudinal balance or
center of gravity of the plesiosaur rather than simply reducing overall buoyancy. In the
case of NJSM 15435, the effect of 6.8 kg of stones as ballast on the buoyancy of a marine
reptile weighing several thousand kilograms is considered to have been negligible.
Summary
Ninety-five gastroliths (6.8 kg) were recovered from within the abdominal
region of a large elasmosaurid plesiosaur (NJSM 15435) in direct contact with matrix
containing whole, fragmentary, and finely ground teleost fish bones, teeth and scales. The
stomach contents of NJSM 15435 indicate a piscivorous diet, with Enchodus and other
small (less than 50 cm / 1.6 feet) clupeomorph fishes being the main food sources. The
location of the gastroliths within the abdominal region of NJSM 15435 and their intimate
association with the fragmentary remains of small fish supports the hypothesis that these
stones aided the plesiosaur in the breakdown of food. Whether or not this was their
primary function or was secondarily related to buoyancy control remains a matter of
debate.
Stomach Contents and Gastroliths
from NJSM 15435 Styxosaurus

|

|

|
Matrix Sample 2 (2 gastroliths) |
Matrix Sample 2 with additional
gastroliths |
Matrix Sample 2- Close up of the
same three gastroliths |

|

|
Three samples of matrix containing
gastroliths and fish bone 'hash' associated with a plesiosaur from the Pierre Shale of
Kansas (scale = mm) |
Matrix Sample 1 (2 gastroliths) |
Matrix Sample 3 (2
gastroliths lying on top of gastralia). A 3rd gastrolith is underneath |
Acknowledgments
We thank Jerome (Pete) Bussen of Wallace, Kansas for donating this unique
specimen to the New Jersey State Museum. This paper would have not been possible without
the collective help of the many volunteers who participated in the digs. The authors also
thank David Parris, Bill Gallagher, and Barbara Grandstaff of the New Jersey State Museum
for the opportunity to prepare and describe the stomach contents, and for their critical
review of the initial manuscript. J. Calcagni provided assistance in preparing the stomach
contents. Charlotte Holton (American Museum of Natural History) provided unpublished
locality data for the Barnum Brown (1904) specimen. L. F. Noè (University of Derby,
Derby, England) provided information on stomach contents and gastroliths associated with
English plesiosaurs. Comments provided by Christian Cicimurri, Philip Currie (Royal
Tyrrell Museum of Paleontology, Earl Manning (Tulane Univ., New Orleans), Glenn Storrs
(University of Cincinnati), and Richard Zakrzewski (Sternberg Museum, Fort Hays State
University) greatly improved upon the final version of the manuscript. Special thanks are
also extended to J. D. Stewart of the Los Angeles County Museum of Natural History for his
identification of the fish remains.
| AUTHORS NOTE: Do not cite or quote
from this webpage. This is a early draft of the published manuscript, with additional or
updated figures. Please refer to: Cicimurri, D. J. and M. J. Everhart, 2001. An elasmosaur
with stomach contents and gastroliths from the Pierre Shale (late Cretaceous) of Kansas.
Kansas Acad. Sci. Trans 104(3-4):129-143. "Feeding Styxosaurus" (above)
is copyright © 2001 by Russell Hawley. |
Literature Cited
Andrews, C. W. 1910. A descriptive catalogue of the marine reptiles of the
Oxford Clay, Part I. British Museum (Nat. Hist.) 1:x-xvii + 205 p.
Bretz, R. F. 1987. Metamorphic, igneous, and sedimentary relationships on the
Sioux Quartzite Ridge, South Dakota, p. 237-242 in S.S. Beuss, ed., Rocky Mountain
Section of the Geol. Soc. Am. Centennial Field Guide (2): 475 pp.
Brown, B. 1904. Stomach stones and the food of
plesiosaurs. Science (New Ser.) 20(501):184-185.
Camp, C. L. 1942. California mosasaurs. Mem. Univ. Calif. 13: vi + 68 p.
Carpenter, K. 1990. Upward continuity of the Niobrara fauna with the Pierre
Shale fauna. Pages 73-81 in Bennett, S. C., ed., Niobrara Chalk Excursion
Guidebook, Univ. Kansas Mus. Nat. Hist. and Kansas Geol. Surv.
Carpenter, K. 1999. Revision of North American elasmosaurs from the Cretaceous
of the Western Interior. Paludicola 2(2):148-173.
Carroll, R. L. 1985. Evolutionary constraints in aquatic diapsid reptiles.
Pages 145-155 in Cope, J. C. W. and P. W. Skelton, eds., Evolutionary case
histories from the fossil record, Spec. Pap. Palæont. 33:202 pp. London, England: The
Palæontological Assoc.
Cicimurri, D. J. and M. J. Everhart. 2001. An elasmosaur
with stomach contents and gastroliths from the Pierre Shale (late Cretaceous) of Kansas.
Kansas Acad. Sci. Trans 104(3-4):129-143.
Cope, E. D. 1868. [Remarks on a new enaliosaurian, Elasmosaurus
platyurus.] Proc. Acad. Nat. Sci. Phil., 20:92-93.
Cope, E. D. 1872. [On a species of Clidastes and on Plesiosaurus gulo
Cope]. Proc. Acad. Nat. Sci. Phil., 24:127-129.
Cott, H. B. 1961. Scientific results of an inquiry into the ecology and
economic status of the Nile Crocodile (Crocodilus niloticus) in Uganda and northern
Rhodesia. Trans. Zool. Soc. London 29(4):212-270.
Currie, P. J. 1981. Hovasaurus boulie: An aquatic eosuchian from the
upper Permian of Madagascar. Palaeontol. Afr. 24:99-168.
Darby, D. G. and R. W. Ojakangas. 1980. Gastroliths from an upper Cretaceous
plesiosaur. Jour. Paleon. 54(3):548-556.
Denton, R. K., J. L. Dobie and D. C. Parris. 1997. The marine crocodilian Hyposaurus
in North America. Pages 375-397 in Callaway, J. M. and E. L. Nichols, eds., Ancient
Marine Reptiles. San Diego.
Elias, M. K. 1931. The geology of Wallace County, Kansas. Kansas Geological
Survey Bulletin 18, 254 pp.
Everhart, M. J. 1999. Evidence of feeding on mosasaurs by the late Cretaceous
lamniform shark, Cretoxyrhina mantelli. Jour. Vert. Paleon. 19(supplement to no.
3):43A-44A, (Abstract).
Everhart, M. J. 2000. Gastroliths associated with
plesiosaur remains in the Sharon Springs Member (late Cretaceous) of the Pierre Shale,
western Kansas. Kansas Acad. of Sci. Trans. 103(1-2):58-69.
Everhart, M. J. 2002. Where the elasmosaurs roam......
Prehistoric Times. 53:24-27.
Everhart, M. J. 2004. Conchoidal fractures preserved on elasmosaur gastroliths
are evidence of use in processing food. Journal of Vertebrate Paleontology 24 (Supplement
to 3): 56A.
Everhart, Michael J. 2005. Oceans of Kansas - A Natural History of the Western
Interior Sea. Indiana University Press, 320 pp.
Everhart, M.J. 2005. Probable plesiosaur gastroliths from the basal Kiowa Shale (Early
Cretaceous) of Kiowa County, Kansas. Kansas Academy of Science, Transactions 108 (3/4):
109-115.
Everhart, M. J. 2005. Elasmosaurid remains from the Pierre Shale (Upper
Cretaceous) of western Kansas. Possible missing elements of the type specimen of Elasmosaurus
platyurus Cope 1868? PalArch 4(3): 19-32. (See also Noè,
L.F. and Gómez–Pérez, M. 2007 (Below).
Everhart, M. J. 2006. The occurrence of elasmosaurids (Reptilia: Plesiosauria) in the
Niobrara Chalk of Western Kansas. Paludicola 5(4):170-183.
Everhart, M. J. 2007. Use of archival photographs to
rediscover the locality of the Holyrood elasmosaur (Ellsworth County, Kansas). Kansas
Academy of Science, Transactions 110(1/2): 135-143.
Gill, J. K., W. A. Cobban, and L. G. Shultz. 1972. Stratigraphy and composition
of the Sharon Springs Member of the Pierre Shale in western Kansas. U. S. Geol. Survey,
Prof. Paper 728, 47 pp.
Gillette, D., 1990. Seismosaurus: The Earth Shaker, Columbia Univ. Press. New
York, 205 pp.
Henderson,
D.M. 2006. Floating point: a computational study of buoyancy, equilibrium, and gastroliths
in plesiosaurs. Lethaia 39:227-244.
Ji, Q., P. J. Currie, M. A. Norell, and Ji S. A. 1998. Two feathered dinosaurs
from northeastern China. Nature, 393:753-761.
Kobayashi, Y., J-C. Lu, Z-M. Dong, R. Barsbold, Y. Azuma, and Y. Tomida. 1999.
Herbivorous diet in an ornithomimid dinosaur. Nature 402:480-481.
Landes, R. W. 1940. Paleontology of the marine formations
of the Montana Group, Pt. 2, of Geology of the southern Alberta plains: Geological Survey
of Canada, Memoir 221, p. 129-217.
Lingham-Soliar, T. 2000. Plesiosaur locomotion: Is the four-wing problem real
or merely an atheoretical exercise? N. Jb. Geol. Palaont. Abh. 217:45-87.
Martill, D. M. 1992. Pliosaur stomach contents from the Oxford Clay. Mercian
Geologist 13(1):37-42.
Martin, J. E. and P. R. Bjork. 1987. Gastric residues associated with a
mosasaur from the late Cretaceous (Campanian) Pierre Shale in South Dakota. Pages 68-72 in
Martin, J. E. and G. E. Ostrander, eds., Papers in Paleontology in Honor of Morton Green,
Dakoterra 3.
Martin, J. E. and L. E. Kennedy. 1988. A plesiosaur with stomach contents from
the late Cretaceous (Campanian) Pierre Shale of South Dakota: A preliminary report. Proc.
South Dakota Acad. Sci. 67:76-79.
Massare, J. A. 1987. Tooth morphology and prey preference of Mesozoic marine
reptiles. Jour. Vert. Paleon. 7(2):121-137.
Matsumoto, T., I. Obata, Y. Okazaki and Y. Kanie. 1982. An interesting
occurrence of a fossil reptile in the Cretaceous of the Obira Area, Hokkaido. Proc. Japan
Acad. B(58):109-113.
Mudge, B. F. 1878. Geology of Kansas. Pages 60-63 in
First Biennial Report of the Kansas State Board of Agriculture. Topeka.
Nicholls, E. L. 1988. Marine vertebrates of the Pembina Member of the Pierre
Shale (Campanian, Upper Cretaceous) of Manitoba and their significance to the biogeography
of the Western Interior Seaway. unpub. doctoral dissertation Univ. Calgary, 317 pp.
Noè,
L.F. and Gómez–Pérez, M. 2007. Postscript
to Everhart, M.J. 2005. Elasmosaurid remains from the Pierre Shale (Upper Cretaceous) of
western Kansas. Possible missing elements of the type specimen of Elasmosaurus
platyurus Cope
1868? In PalArch’s Journal of Vertebrate
Palaeontology 4, 3: 19–32. PalArch’s
Journal of Vertebrate Palaeontology 2(1): 1-9
Parrish, J. T. and D. L. Gaultier. 1993. Sharon Springs Member of the Pierre
Shale: upwelling in the Western Interior Seaway? Pages 319-322 in Caldwell, W. G.
E. and E. G. Kauffman, eds., Evolution of the Western Interior Basin. Geol. Assoc. Canada
Special Paper 39.
Pollard, J. E. 1968. The gastric contents of an ichthyosaur from the lower Lias
of Lyme Regis, Dorsett. Paleontology 11(3):376-388.
Reiss, J. and E. Frey. 1991. The evolution of underwater flight and the
locomotion of plesiosaurs. Pages 131-144 in Rayner, J. M. V., and R. J. Wooten,
eds., Biomechanics of Evolution. Cambridge Univ. Press, Cambridge.
Riggs, E. S. 1939. A specimen of Elasmosaurus serpentinus. Field Mus.
Nat. Hist., Geol. Survey, Report of Investigations 6(5):385-392.
Sander, P. M., O. C. Rieppel and H. Bucher. 1997. A new pistosaurid (Reptilia:
Sauropterygia) from the Middle Triassic of Nevada and its implications for the origin of
the plesiosaurs. Jour. Vert. Paleon. 17(3):526-533.
Sato, T. and K. Tanabe. 1998. Cretaceous plesiosaurs ate ammonites. Nature
394:629-630.
Sato, T. and G. W. Storrs. 2000. An early polycotylid plesiosaur (Reptilia:
Sauropterygia) from the Cretaceous of Hokkaido, Japan. Jour. Paleon. 74(5):907-914.
Schäfer, W., 1972. Ecology and Palaeoecology of Marine Environments. C.Y.
Craig, ed., The University of Chicago Press, Chicago, 568 pp.
Schmeisser,
R.L. and Gillette, D.D. 2009. Unusual occurrence
of gastroliths in a polycotylid plesiosaur from the Upper Cretaceous Tropic Shale,
southern Utah. PALAIOS 2009 24: 453-459.
Schwimmer, D. R. 1997. Late Cretaceous dinosaurs in the eastern USA: a
taphonomic and biogeographic model of occurrences. Dinofest International Symposium
Proceedings, Acad. of Natural Sciences:203-211.
Schwimmer, D. R., J. D. Stewart and G. D. Williams. 1997. Scavenging by sharks of
the genus Squalicorax in the late Cretaceous of North America. PALAIOS
12:71-83.
Seeley, H. G. 1877. On Mauisaurus gardneri Seeley, an elasmosaurian from
the base of the Gault of Folkestone. Quart. Jour. Geol. Soc. London, 33:541-546.
Shimada, K. 1997. Paleoecological relationships of the late Cretaceous
lamniform shark, Cretoxyrhina mantelli (Agassiz). Jour. Paleon. 71(5):926-933.
Shuler, E. W. 1950. A new elasmosaur from the Eagle Ford Shale of Texas.
Fondren Science Series 1(2):1-33.
Storrs, G. W. 1981. A review of occurrences of the Plesiosauria (Reptila:
Sauropterygia) in Texas, with description of new material. Masters Thesis, The University
of Texas at Austin, 226 pp.
Storrs, G. W. 1993. Function and phylogeny in sauropterygian (Diapsida)
evolution. Am. Jour.Sci. 293A:63-90.
Storrs, G. W. 1999. An examination of Plesiosauria (Diapsida: Sauropterygia)
from the Niobrara Chalk (upper Cretaceous) of central North America. Univ. Kansas Paleont.
Cont. (New Ser.), (11):15 pp.
Tarlo, L. B. 1959. Pliosaurus brachyspondylus Owen from the Kimmeridge
Clay. Palæontology 1:283-291.
Taylor, G., 1999. Winging it. New Scientist 163(2201):28-32.
Taylor, M. A. 1981. Plesiosaurs - rigging and ballasting. Nature 290:628-629.
Taylor, M. A. 1993. Stomach stones for feeding or buoyancy? The occurrence and
function of gastroliths in marine tetrapods. Phil. Trans. Roy. Soc. London, B 341:163-175.
Taylor, M. A. 1994. Stone, bone or blubber? Buoyancy control strategies in
aquatic tetrapods. Pages 205-229 in Maddock, L., Q. Bone, and J. M. V. Rayner,
eds., Mechanics and physiology of animal swimming. Cambridge Univ. Press, Cambridge.
Taylor, M. A., D. B. Norman, and A. R. I. Cruickshank. 1993. Remains of an
ornithischian dinosaur in a pliosaur from the Kimmeridgian of England. Palæontology
36(2):357-360.
Thompson, W. A.,
Martin, J. E. and Reguero, M. 2007. Comparison of
gastroliths within plesiosaurs (Elasmosauridae) from the Late Cretaceous marine deposits
of Vega Island, Antarctic Peninsula, and the Missouri River area, South Dakota. Pages
147-153 in Martin, J. E. and Parris D. C. (eds.), The
Geology and Paleontology of the Late Cretaceous Marine Deposits of the Dakotas. Geological
Society of America, Special Paper 427.
Wahl, W. 1998. Plesiosaur gastric contents from the upper Redwater Shale (lower
Oxfordian) of the Sundance Formation (Jurassic) of Wyoming. Jour. Vert. Paleon. 18
(Supplement to No. 3):84A, (Abstract).
Welles, S. P. and J. D. Bump. 1949. Alzadasaurus pembertoni, a new
elasmosaur from the Upper Cretaceous of South Dakota. Jour. Paleon. 23(5):521-535.
Whittle, C. H. and M. J. Everhart. 2000. Apparent and implied evolutionary
trends in lithophagic vertebrates from New Mexico and elsewhere. Pages 75-82 in
Lucas, S. G. and A. B. Heckert, eds., Dinosaurs of New Mexico, New Mexico Museum of
Natural History and Science, Bulletin 17, 230 pp.
Wiffen, J. and W. L. Moisley. 1986. Late Cretaceous reptiles (families
Elasmosauridae and Pliosauridae) from the Mangahouanga Stream, North Island, New Zealand.
New Zealand Journ. Geol. Geophys. 29:205-252.
Williston, S. W. 1893. An interesting food habit of
the plesiosaurs. Kansas Acad. Sci. Trans. 13(2):121-122.
Williston, S. W. 1904. The stomach stones of the plesiosaurs. Science (New
Ser.) 20:565.
Williston, S. W. 1914. Water reptiles of the past and present. Univ. of Chicago
Press. 251 pp.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}