 |
Where the elasmosaurs roam: Separating
fact from fiction
Original article published as: Everhart, M. J. 2002. Where the
Elasmosaurs roam......
Prehistoric Times 53:24-27
Copyright © 2002-2009 by Mike Everhart; Last revised
09/18/2009
LEFT: Elasmosaurus adapted from a painting by Doug Henderson. Copyright ©
Doug Henderson; used with permission of Doug
Henderson |
Read more about KANSAS
PLESIOSAURS
The warm, silt-laden water felt strange to the young, long-necked
plesiosaur as he slowly worked his way upstream against the strong current of the river.
His streamlined body seemed less buoyant in the fresh water and that made raising his head
above water to breathe a bit more difficult. Unlike the ocean that was his home, he could
not see very far in the murky water, so he moved cautiously along the sandy bottom, trying
to stay in the deeper reaches of the main channel. Dimly he remembered being here several
times before with a much larger member of his species. At the time, it was all he could do
to stay close to her body and not be swept away by the current. Now sensory cells in his
nasal passages ‘tasted’ strange new smells as the water passed in through his
nose and exited his slightly opened mouth. Although he was hungry after his long journey,
his instincts drove him up this river for something else.
Within a mile from where the river entered the ocean, the sand bottom of
the river had turned to coarser gravel. The plesiosaur lowered his head and began to probe
the gravel with his lower jaw. Clouds of silt and other debris rose wherever his chin
touched the bottom. The sensations received through his lower jaw told him that this was
not what he was looking for and he began swimming again. Holding his head and neck stiffly
in front of his body, he relied on alternating beats of his wing-shaped front and rear
flippers to provide the thrust that he needed to ‘fly’ forward through the
water.
The river narrowed as he swam upstream and the current became faster.
Swimming against a current was not something that he was used to doing. Such currents were
hardly ever found in the calm, shallow ocean where he had spent almost all of his life.
Within a short distance, he began to encounter large rocks that formed eddies in the
current. It was behind a rock larger than himself that he finally found what he was
looking for. Small round stones had collected on the downstream side of the boulder.
Twisting his head slightly to the side, he was able to see that there were many more here
than he would need. Slowly and delicately, he picked up one smooth rock after the other
with his long, slender teeth. With each, he raised his head slightly and swallowed,
feeling the hard stone move into his throat and start the slow trip down his 15 foot long
esophagus to his stomach. He continued selecting and swallowing the round stones until he
had satisfied his need. Raising his head above the surface briefly to breathe, his sharp
underwater vision saw only a dark green blur that was the dense forest that crowded over
the riverbank. Again he submerged, then turned and began his return journey back to the
sea.
His movement flushed a small fish from its hiding place near the large
rock. Before the fish could flee, the plesiosaur reacted and trapped it between jaws
filled with long, interlocking teeth. Raising his head just above the surface, he
swallowed his prey headfirst, then exhaled again and refilled his lungs. Vaguely
satisfied, he submerged his head and began to swim downstream with the current with slow,
deep strokes of his paddles.
 |
Although isolated fragments of plesiosaurs had been described from
North America earlier in the 19th Century by Joseph Leidy and others, the
remains discovered by Dr. Theophilus H. Turner (1841-1869) near Fort Wallace in western
Kansas in 1867 represented the first nearly complete specimen and the first known
elasmosaur. Dr. Turner was an Army surgeon assigned to Fort Wallace. He found the remains
of the huge animal while exploring exposures of the Pierre Shale along the right-of-way of
the approaching Union Pacific Railroad near McAllaster Butte in present-day Logan County,
Kansas. Dr. Turner gave two of the
vertebrae of his "extinct monster" to John LeConte to take back for further
examination in "eastern scientific institution." At the time, Dr. LeConte was a
member of the group surveying an extension of the Union Pacific Railway. Upon completion
of the survey, he returned to Philadelphia and took the vertebrae to E. D. Cope at the Academy of
Natural Sciences in Philadelphia. |
| Cope immediately recognized the bones as belonging to a large
plesiosaur and wrote back to Dr. Turner about the possibility of obtaining the rest of the
specimen, offering to pay all expenses. Turner agreed to ship the rest of the specimen to
Cope and, with a party of other soldiers from Fort Wallace, began digging on it in early
December, 1867. Work progressed rapidly and Turner was able to ship several hundred pounds
of bones and matrix by rail in early 1868. |
After receiving the shipment in March, 1868, Cope hurriedly assembled the bones
into a strange and hereto unknown extinct animal which he named Elasmosaurus platyurus
or roughly translated, "Flat-tailed plate-reptile." As was an accepted method of
the times, Cope rushed the news of the discovery and a description into publication in a
popular scientific journal without review by his peers. Unfortunately, in his haste, and
following some of the early work on American plesiosaurs by his mentor, Joseph Leidy, Cope
had placed the head of the plesiosaur on the wrong end. Contrary to popular belief,
however, it was not Cope’s rival, O. C. Marsh, who discovered the mistake. It was
Joseph Leidy himself, who announced two years later at another meeting of the Academy that
Cope’s reconstruction was wrong and that his own, earlier work on plesiosaurs was
also in error. Cope tried to minimize the damage by offering to buy back all copies of the
journal but it was too late. In spite of Cope’s efforts and the immediate publication
of a partially corrected description, O. C. Marsh would later make
good use of the opportunity to embarrass his rival. This incident was probably the opening
battle in the 19th Century American paleontology conflict that later became
known as "The Bone Wars".
The first nearly complete plesiosaur was discovered in the Jurassic rocks of
Lyme Regis, England by Mary Anning in the winter of 1820-21. The name was given by the
Reverend William Conybeare and means "near-reptile", a reference to the view
that plesiosaurs were closer to reptiles than were the ichthyosaurs. Several complete
specimens were already in European museums by the time that the American West was being
settled. Some of these plesiosaurs like Plesiosaurus and Cryptoclidus had
relatively long necks and small heads, and looked very much like their Cretaceous cousins.
However, there is some evidence that many of these early plesiosaur lineages became
extinct at the end of the Jurassic. Whatever the case, the numbers and diversity of
plesiosaurs appears to have declined steadily through the Cretaceous.
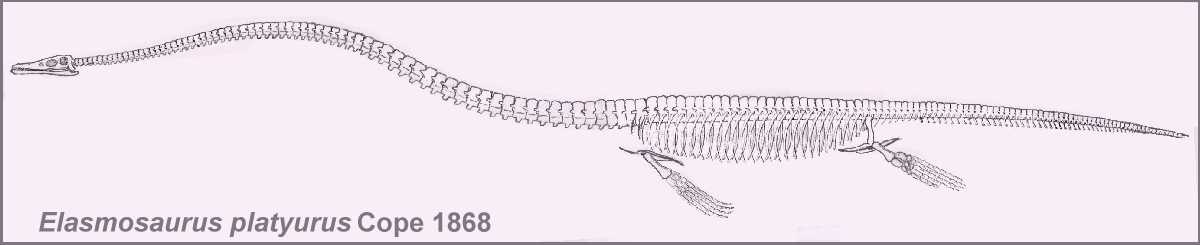
Elasmosaurus platyurus
was the first of several species of the very long-necked plesiosaurs (elasmosaurs) to be
discovered and described from the late Cretaceous deposits in western Kansas, the Midwest
and elsewhere in North America. Eventually, elasmosaurs would be found in Cretaceous
marine deposits of almost every continent, including Australia and New Zealand, and even
Antarctica. From where most of their fossils have been found, it appears that they
preferred cooler waters found at higher latitudes to those of warmer equatorial climates.
In North America, they are found more often in the Cretaceous deposits of Canada than in
the central United States, and are extremely rare in the Gulf area.
Plesiosaurs are fascinating creatures, both for what we know and don’t
know about them. By the Late Cretaceous, they had evolved into two distinct, but closely
related groups that are generally referred to as short-necked (polycotylids) and
long-necked (elasmosaurs). The short-necked variety were fairly small, reaching lengths of
no more than 12-15 feet. They had long narrow jaws filled with slender, seizing teeth. It
is likely that they were capable of swimming rapidly in pursuit of the small fish and
cephalopods that were their prey. Elasmosaurs, on the other hand, had somewhat broader but
much shorter heads on extremely long necks. They reached lengths of 45 feet or so, and
weighed several tons as adults. They were probably slow moving, and not much of a threat
to anything larger than a small fish. Their size, weight, and highly
modified limbs probably meant that they could not have crawled ashore to lay eggs.
For various reasons, elasmosaurs seem to have captured the imagination of
almost anyone interested in ancient creatures. In spite of their extinction more than 65
million years ago, their mystique lives on in Nessie, Champ and other "sea- and
lake-serpents" around the world, basking shark carcasses netted by Japanese fishing
boats and other unexplained sightings. All that really remains of them, however, are their
bones. In the following text, I will try to put some flesh on these old bones.
Plesiosaurs have been described as "a snake drawn through the shell of a
turtle"(*). While this is an exaggeration of sorts, the tear-drop shaped body of an
elasmosaur was almost totally enclosed by long ribs, large pectoral and pelvic girdles and
a continuous series of gastralia across the belly. A long neck was attached to the front
and four wing-like paddles projected at nearly right angles. In one species (Elasmosaurus
platyurus), the neck had seventy or more vertebrae and was more than half the length
of the body. Since their discovery, this unusually long neck has produced a number of
incorrect descriptions of its "snake-like" flexibility.
| (*) While the source of this quote has been a mystery for a
number of years, it was partially solved by Richard
Forrest in September, 2005. Richard found that it was apparently first used in a
German publication (Dames,
Wilhelm [Barnim]. 1895. Die Plesiosaurier der
süddeutschen Liasformation. Abhandlungen
der Königlichen Akademie der Wissenschaften in Berlin
(physics-math), pp.
1-83, pls. i-v. [The plesiosaurs of the Lias
Formation of southern Germany]). Quote "...dass man Plesiosaurus mit
einer durch einen Schildkrõtenpanzer gezogenen Schlange vegleich ..." which
translates as "...one can compare the Plesiosaurus to a snake threaded through the
shell of a turtle.." It is also mentioned in Zarnik (1925), which was written in
Serbo-Croat (Zarnik, B.; 1925; Sketologiji plesiosauriija, sa prinosima mehanici
kraljeznice u recentnih sauropsida.; Societas Scientiarum Naturalium Croatica, Hrvatskoga
Naravoslovno Drestva; 38-39 pp.424-473 (On the ethology of plesiosaurs with contributions
to the mechanism of the cervical vertebrae of recent sauropsids). See the latest on the search for the origin of this quote here.
For more information on Jurassic plesiosaurs: Adam
Smith's The Plesiosaur Directory
|

|
| E. D. Cope, in his 1872 paper "On the geology and
paleontology of the Cretaceous strata of Kansas," was certainly among the first and
most influential when he mused about the appearance of swimming plesiosaurs. He wrote,
''Far out on the expanse of this ancient sea might be seen a huge, snake-like form which
rose above the surface and stood erect, with tapering throat and arrow-shaped head, or
swayed about describing a circle of twenty feet radius above the water. ... Should several
have appeared together, we can easily imagine tall, flexible forms rising to a height of
the masts of a fishing-fleet, or like snakes, twisting and knotting themselves
together." Later, he added, the "snakelike neck was raised high in the air, or
depressed at the will of the animal, now arched swanlike, preparatory to a plunge after a
fish, now stretched in repose on the water, or deflexed in exploring the depths
below." |
Many paleo-artists, including Charles Knight (Above), have drawn popular but
anatomically impossible representations of elasmosaurs based on these early, fanciful
descriptions. These ideas still persist in spite of publications by Williston (1914) and
others that have shown that the cervical vertebrae of elasmosaurs had limited movement up
and down and only slightly more in a horizontal plane There are good reasons for this
limited flexibility which are discussed below.

|
| Another similar misconception about the long neck of plesiosaurs
is shown in pictures of elasmosaurs cruising on the surface with their head and neck
arched swan-like, high above the water as they searched for prey. Unless the laws of
physics were suspended on the behalf of these extinct creatures, it would have been
impossible for them to lift much more than their head above water to breathe. If you would
like to prove this for yourself, try lifting something long and heavy from one end while
floating in water and not touching the bottom. As you try to raise the object, your feet
and the lower part of your body will also begin to surface to counter-balance any weight
above the water. In elasmosaurs, the weight of the long neck (as much as a ton or more in
an adult animal) dictated that the center of gravity was just behind the front flippers.
Holding the head and neck above the surface was something that could never happen unless
the rest of the body was setting on a firm bottom in shallow water (a possibly fatal
situation for the elasmosaur!). Even then, the limited movement between the vertebrae,
limited musculature, and the sheer weight of the neck would severely limit the height to
which the head could have been raised. |
Still another problem that becomes evident with the long neck of elasmosaurs is
what happens when the head is moved to one side or the other while the animal is moving.
Sitting twenty feet or more in front of the flippers, the head (and neck) would act as a
highly effective rudder whenever it was moved away from the longitudinal axis of the
animal. In other words, moving the head to the right while swimming would cause the whole
body of the plesiosaur to turn in that direction. While it is possible that plesiosaurs
used their head as a rudder to change directions while swimming, it would also be nearly
impossible for them to swim in a straight line with their heads darting from side to side,
or up and down, in search of prey as shown in many reconstructions. In any case, it seems
likely that the tiny brain of an elasmosaur would have been very busy trying to maintain
the position of the head during stationary feeding activities through fine movements of
the large paddles located up to twenty-five and thirty-five feet to the rear. No
disrespect for the mental capacity of plesiosaurs is intended here since plesiosaurs, and
elasmosaurs in particular, were very successful predators for millions of years, even if
we brainy humans don’t yet understand how they did it.
 |
Thalassomedon
haningtoni from the late Cretaceous (Turonian) Graneros Shale of Colorado and
Nebraska, and Elasmosaurus
platyurus from the Campanian Pierre Shale of Kansas were the longest of the
elasmosaurs presently known: probably up to 14 meters or 45 feet in length. The length of
the neck of elasmosaurs appears to have increased through time, conveying some adaptive
advantage for this group that we do not fully understand. While other large marine
predators of the time, including their short-necked, polycotylid
cousins, snake-like mosasaurs, giant fish such as Xiphactinus,
and huge Ginsu sharks (Cretoxyrhina mantelli), used speed
to chase down or ambush their prey, it is more likely that elasmosaurs used stealth to
stalk the schools of small fish that were their primary food source. A long neck would
allow the large body of the plesiosaur to remain concealed far below a school of fish
while the small head moved slowly at the end of its long neck to approach the unlucky
victim from below. Having their eyes on top of
their head, directed generally forward and upward, as is the case in most elasmosaurs,
seems to support this method of feeding. The eyes were not unusually large but may have
been capable of stereoscopic vision, something that would be useful for accurately
locating small prey. It is likely that the head was moved fairly close to the prey before
striking. Precise and rapid coordination between the eyes and the elasmosaurs paddles
would have been essential for successful hunting of highly mobile fish and other marine
life. Hunting from below may have also been used as a strategy to silhouette the prey of
the elasmosaur against the sunlit surface while using the darker, deeper waters for
concealment. |
We know very little about the other senses of elasmosaurs. Some authors have
suggested that the nostrils of plesiosaurs were well positioned to passively direct a
continuous flow of water through the nasal cavity where the senses of taste and smell may
have been useful in detecting prey. Williston notes that the inner ear (semi-circular
canals) of plesiosaurs was large and well developed. This area serves to maintain
equilibrium and coordinate muscular control and infers that plesiosaurs were probably
quite graceful in their movements. While there is no evidence of external ears in
plesiosaurs, they may have evolved a sensory line system that was used to locate prey from
the vibrations generated by their movement. It appears likely, however, that they relied
primarily on their eyesight to find food.
Evidence from stomach contents preserved in plesiosaur remains indicates that
the prey that they ate was limited to small sized fish and invertebrates. Earlier
plesiosaurs from the Jurassic and early Cretaceous appear to have preferred squid and
other cephalopods, while the later and longer necked elasmosaurs fed on small fish such as
Enchodus. While elasmosaurs were huge by any measure, their heads remained
relatively small in relation to the size of their bodies A forty foot elasmosaur had a
skull length of about 20 inches or about 4% of the total length of the animal. Unlike
those of most mosasaurs, plesiosaur skulls were rigidly constructed. This meant that the
cross-sectional size of the prey they could swallow was limited by the fixed distance
between the hinge points (quadrates) of their jaws at the back of the skull. Even in
the largest of the elasmosaurs this distance was no more than about 6-8 inches, and the
prey that they fed on was probably no longer than about 18 inches in length. Their
slender, interlocking teeth also appear to argue for trapping small prey. In all cases,
plesiosaur teeth appear to have been used to seize small fish or invertebrates, and not to
tear flesh or otherwise dismember larger prey. Once seized, the prey would have to be
swallowed whole.
Because of their unusual body-plan, elasmosaurs were most likely slow moving
animals, using their paddles as wings or foils to generate the lift necessary to
‘fly’ through the water. This type of swimming is more efficient than the rowing
or paddling (i.e., a duck, Grebe or other diving bird) methods that are sometimes
depicted. While modern underwater "fliers" such as penguins use only their front
limbs, most researchers now believe that plesiosaurs used a coordinated movement of both
pairs of limbs. Others think that the hind limbs did not have either the range of movement
or the musculature necessary to effectively generate the lifting forces necessary for this
method of swimming. Since the hind limbs are located well behind the center of gravity,
they may have been more effectively used for steering. In that case, the rear limbs may
have been rudders, paddles and/or stabilizers, especially to control the position of the
body and head while remaining relatively motionless in the water during feeding.
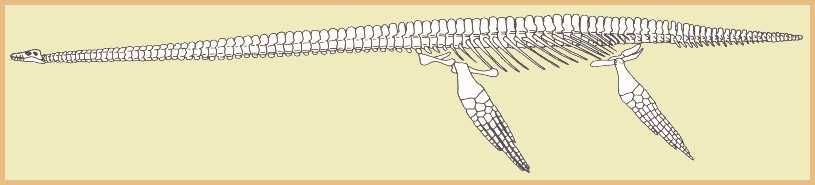
|
| In plesiosaurs, both pairs of limbs are about equal in size and
are highly modified into a rigid paddle. The upper end of the paddle, called the propodial
(humerus / femur) is attached to the body at the shoulders and hips. The lower limb bones
(radius/ulna and tibia/fibula) are reduced to flat, polygonal shapes that interlock with
the smaller but similar shaped bones of the wrist and ankle. |
The finger and toe bones (phalanges and tarsals) were hour-glass shaped. They
fitted closely together with the bones of the adjoining fingers and were probably held
tightly within a sheath of muscles, tendons and ligaments. Although plesiosaurs have five
‘fingers’, they exhibit a condition called hyperphalangy which means that they
have many more individual finger bones than usual (humans have 4 bones in each finger;
plesiosaurs had as many as 24!). The net effect of the compacted lower arm / leg and wrist
/ ankle structures, and the extra finger / toe bones was to modify what had been a walking
limb into a long, tapering paddle that had limited flexibility. In cross-section, the
distal portion of the limb was thicker on the leading edge than the trailing edge. It had
literally become a wing or hydrofoil for flying through the water. Movement of the paddles
in a manner similar to that of a bird’s wing in flight created lift and pulled the
animal through the water. Unlike birds, however, the wrist was ‘locked’ and the
paddles could not be folded or otherwise flexed.
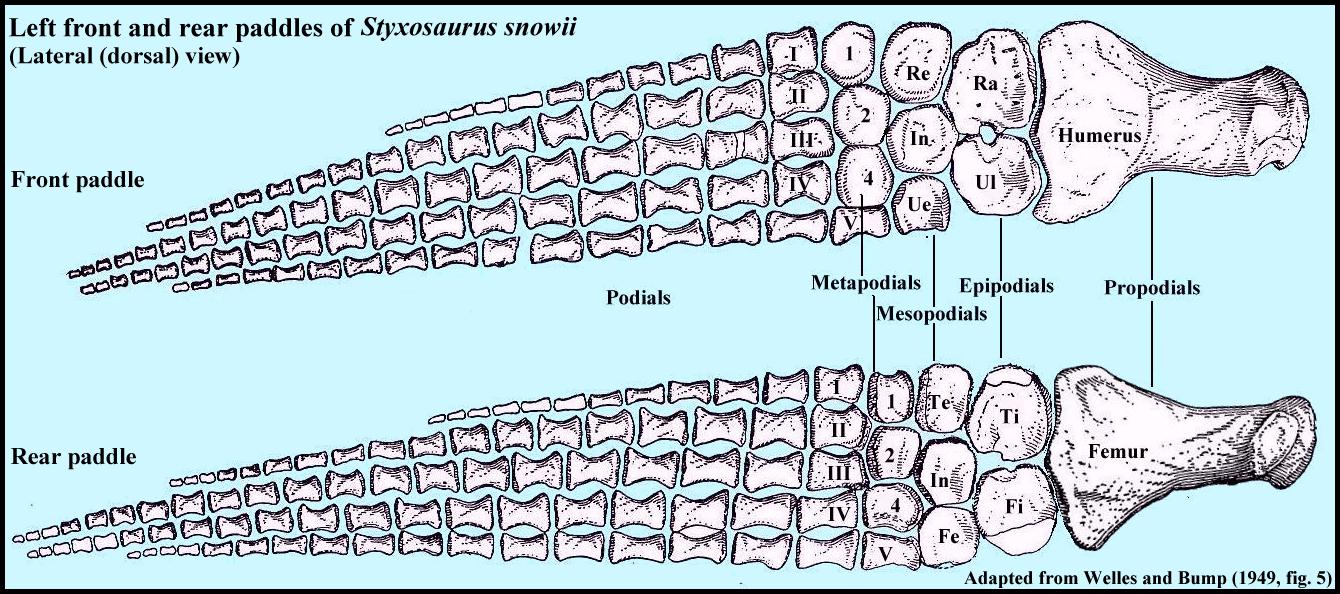
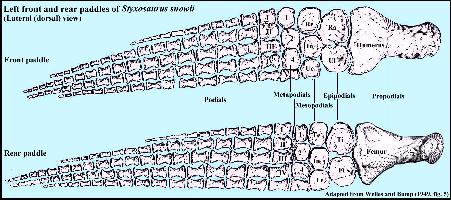 |
Although plesiosaurs evolved from terrestrial reptiles with four
legs, the limbs were highly modified into paddles, and lost most of the mobility of the
elbow / knee, wrist / ankle and fingers / toes. The bones of the wrist became specialized
to the point that they have had to be given different names (no longer carpals or
tarsals). LEFT: A lateral (dorsal) view of the left front and rear
limbs of Styxosaurus snowii (SDSMT 451) as adapted from
Welles and Bump (1949, pl. 5) Abbreviations: Ra = Radius; Ul = Ulna; Ti =
Tibia; Fi = Fibula; Re = Radiale; In = Intermedium; Ue = Ulnare; Te = Tibiale; Fe =
Fibulare. |
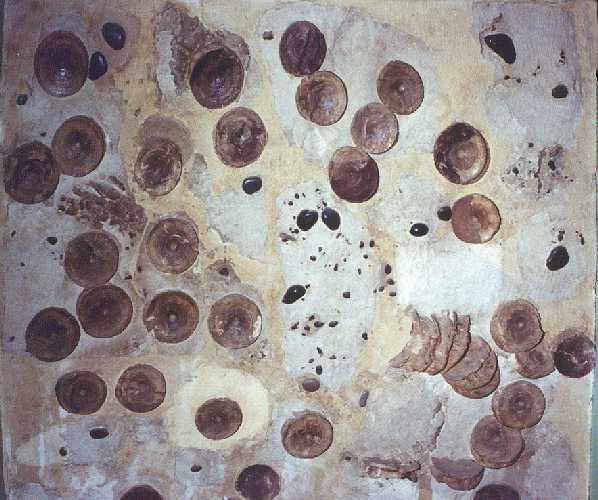
One other peculiarity that has been noted among long necked plesiosaurs since
their discovery are the masses of gastroliths or stomach stones
that are commonly found associated with their remains. These rounded and sometimes highly
polished rocks are usually located inside the abdomen of complete specimens. There have
been as many as several hundred stones of various sizes, from egg-sized down to pea-sized
or smaller found associated with plesiosaurs. In the case of a recently discovered
specimen from Antarctica, there were thousands of small quartzite pebbles (half-inch or
less) in the stomach area. Typically, most gastroliths are hard rocks such as quartz,
quartzite, or chert. Occasionally they are granite, basalt or other volcanics. Until
recently, the largest gastroliths have been about as big as a tennis ball, but in two
Kansas specimens found in the early 1990s, they were much larger. The largest stones in a
University of Kansas specimen weighed about three pounds each and were nearly 4 inches
in diameter (softball size!). These are the largest individual sizes and the greatest
total weight of gastroliths ever documented from a plesiosaur or from any animal. The
total weight of the stomach stones from this plesiosaur was about 30 pounds but
considering the weight of the animal (about 3 tons), their relative weight is rather
insignificant and they probably represent much less than weight of the daily food intake.
More commonly, the total weight of gastroliths documented from individual plesiosaurs is
less than ten pounds. It appears highly unlikely that these stones would have been
effective in changing the buoyancy of large animals that had to consume a lot of food
daily in order to sustain themselves while living in mid-ocean. Even the buoyancy changes
caused by inhalation and exhalation through a 25-foot long trachea would have been more
than could have been offset by the small amount of weight represented by the gastroliths.
This leads us to the question of what gastroliths were actually used for? While
early fossil collectors such as Benjamin Mudge concluded from
plesiosaurs found in Kansas in the 1870s that the stomach stones were used as "an aid
to digestion similar to many living reptiles and birds," Samuel Williston and others
initially believed that they were swallowed for other reasons, including a food craving or
being mistaken for prey. Currently, most researchers believe that they were used in some
manner for buoyancy control or for adjusting longitudinal balance. However, a nearly
complete specimen of Styxosaurus snowii recently
discovered in the Pierre Shale of western Kansas provides a rather convincing
counter-argument. These remains included about a hundred gastroliths, fish bones, scales
and teeth co-mingled in the stomach area. Most of the fish parts appeared to have been
ground up into very small (0.1 in. or less) fragments. This suggests that whatever use
they may have had for buoyancy control, they were definitely useful as an aid to
digestion.
So where did these gastroliths come from? Mudge noted that the Kansas
gastroliths "were the more curious, as we never found such pebbles in the chalk or
shales of the Niobrara." Williston noted that some of the quartzite stones were
similar to boulders found in northwestern Iowa or in the Black Hills of South Dakota. Both
of these potential sources are 400 to 500 miles from where the plesiosaur remains that
contained them were found. Whatever their origin, they certainly did not come from the
soft mud bottom of the Western Interior Sea that covered Kansas. This in itself implies
that the plesiosaurs had to travel long distances to the sources of these stones. The fact
the stones are always rounded and smooth also means that the stones were eventually worn
down to sizes small enough to pass though from the crop or stomach into the gut and had to
be replaced periodically throughout the life of the plesiosaur.
A final subject is the issue of plesiosaurs crawling ashore to lay their eggs.
It has been long assumed that because they are reptiles, they must have laid eggs.
Consequently, plesiosaurs sunning
themselves on the beach have been the subject of imaginative works by paleo-artists
for more than 150 years. While almost all reptiles do lay eggs, it is highly unlikely that
large marine reptiles could have reproduced in that manner. Turtles appear to be the only
group of marine reptiles that still come ashore to lay their eggs. Unlike turtles, the
limbs of plesiosaurs were so modified for use in underwater flying that they would have
been too rigid to be used effectively for movement on land, much less for scooping out a
nest for eggs in beach sand. In addition, the length and weight of an elasmosaur’s
long neck would have effectively counter balanced the back half of the animal, lifting it
off the ground and probably making the rear flippers useless for movement on land, or
anything else, under the best of circumstances.
Ichthyosaurs have long been known to have given live birth to their young as
evidenced by well preserved remains in the Holzmaden Shale of Germany. A mosasaur (Plioplatecarpus) specimen found recently in the Pierre
Shale of South Dakota included the remains of several young of the same species in the
pelvic region. There is also at least one specimen of a short-necked plesiosaur (Dolichorhynchops osborni)
known with fetal material in the abdomen. It appears likely that live birth is one of the
necessary adaptations that egg-laying reptiles had to make in order to successfully return
to life in the ocean. If so, this raises additional questions as to how marine reptiles
nurtured a fetus inside the mother’s body for a relatively long period of gestation.
Did they simply retain the developing embryos and yolk sacs inside the mother’s body
until they were ready to hatch as do some modern snakes? Or did had they developed some
other method of nurturing baby plesiosaurs? We may never know. It is apparent that
similarly-sized dinosaurs laid relatively small eggs which were limited in size by the
ability of the porous shell to exchange oxygen and carbon dioxide for the developing fetus
while supporting the weight of the egg. Plesiosaurs were highly successful animals that
clearly had to have evolved some efficient method for reproduction in the marine
environment.
Giving live birth to their young probably also meant that they provided some
form of parental care. Turtles and other reptiles which lay many eggs at one time depend
on safety in numbers for survival of a few of their young. Animals which invest their
energy in birthing a few larger babies generally provide some sort of protection for those
young as a means of improving their chances to survive to adulthood. Like some dinosaurs,
plesiosaurs may have traveled together in small groups for that purpose. The survival rate
for small, young animals in the middle of an ocean populated with giant predatory fish,
Great White-sized Ginsu (Cretoxyrhina) sharks, and 30 to 50 foot long, hungry
mosasaurs would have otherwise been close to zero. As it was, plesiosaurs may have been
driven to the edge of extinction by the "mosasaur explosion" during the late
Cretaceous. We do know from the bite marks on plesiosaur bones that their carcasses were
scavenged by sharks, and that readily detachable parts, such a limbs, tails, heads and
necks, were often carried away. The remains of a very large Cretoxyrhina shark at
the University of Kansas includes more than a hundred polished
gastroliths that most likely were the result of the shark scavenging on the carcass of
a plesiosaur.
For all of their size and amazing adaptations to life in the ocean, it appears
that plesiosaurs were an evolutionary dead end and were on their way out when the
Cretaceous Period ended. Their greatest diversity and numbers probably occurred during the
Jurassic or early Cretaceous. By early Maastrichtian time (78 mya) there were only a few
species each of elasmosaurs and short-necked plesiosaurs left in Earth’s oceans. Like
the ichthyosaur fish-lizards that became extinct in the early Cretaceous, plesiosaurs were
probably losers of an evolutionary arms race for the same prey that first saw the rise of
larger, faster bony fish and sharks as competitors during the early Cretaceous and then
the explosive entry of the highly adaptable mosasaurs in the Late Cretaceous. Not only
were they probably out-completed for food by larger and more aggressive mosasaurs, in at
least one known case, they were also mosasaur food.
References and further reading:
Almy, K. J. 1987. Thof’s dragon and the letters of Capt.
Theophilus Turner, M.D., U.S. Army, Kansas History Magazine, 10(3):170-200.
(Background on the discovery of Elasmosaurus platyurus,
Cope’s "head on the wrong end" plesiosaur - historical)
Brown, B. 1904. Stomach stones and the food of
plesiosaurs. Science, 20(501):184-185. (Gastroliths mixed with stomach contents in
plesiosaurs)
Carpenter, K. 1994. Comparative cranial anatomy of two North American
plesiosaurs (Reptilia: Sauropterygia) and a review of some North American elasmosaurs,
unpublished draft.
Carpenter, K. 1994, A review of some short-necked plesiosaurs from the
Cretaceous of North America, unpublished draft.
Carpenter, K. 1996. A review of short-necked plesiosaurs from the
Cretaceous of the Western Interior, North America, N. Jb. Geol. Palaont. Abh.,
(Stuttgart), 201(2):259-287.
Carpenter, K. 1997. Comparative cranial anatomy of two North American
Cretaceous plesiosaurs, In Calloway J. M. and Nicholls, E. L., eds, Ancient
Marine Reptiles, Academic Press, pp. 191-216
Carpenter, K. 1999. Revision of North American Elasmosaurs from the Cretaceous
of the Western Interior, Paludicola, 2(2):148-173.
Cicimurri, D. J. and M. J. Everhart. 2001. An elasmosaur with stomach contents and
gastroliths from the Pierre Shale (late Cretaceous) of
Kansas.
Kansas Academy of Science, Transactions 104(3-4):129-143.
Cope, E. D. 1868. Remarks on a new enaliosaurian, Elasmosaurus
platyurus. Proceedings of the Academy of Natural Sciences of Philadelphia
20:92-93. (for meeting of March 24, 1868)
Cope, E. D. 1868. (on new species of extinct reptiles). Proceedings of the
Academy of Natural Sciences of Philadelphia 20:181. (for meeting of July 14, 1868.
Cope, E. D. 1868. On some Cretaceous reptilia. Proceedings of the Academy of
Natural Sciences of Philadelphia 20:233-242. (for meeting of Oct. 27, 1868)
Cope, E. D. 1868. (A resolution thanking Dr. Theophilus
Turner for his donation of the skeleton of Elasmosaurus platyurus). Proceedings
of the Academy of Natural Sciences of Philadelphia 20:314
.
Cope, E. D. 1868. On the genus Laelaps. American Journal of Science
ser. 2, 46(138):415-417.
Cope, E. D. 1870. On Elasmosaurus platyurus
Cope. American Journal of Science ser. 2, 50(148):140-141.
Cope, E. D. 1870. Additional note on Elasmosaurus.
American Journal of Science ser. 2, 50(149):268-269.
Cope, E. D. 1870. Extinct Batrachia, Reptilia and Aves of North
America. Transactions of the American Philosophical Society. New Series 14:1-253.
Darby, D. G. and R. W. Ojakangas. 1980. Gastroliths from an Upper
Cretaceous plesiosaur, Journal of Paleontology, 54(3):548-556.
Everhart, M. J. 2000. Gastroliths
associated with plesiosaur remains in the Sharon Springs Member of the Pierre Shale (late
Cretaceous), Western Kansas. Kansas Academy of Science, Transactions 103(1-2):58-69.
Everhart, M. J. 2003. First records of plesiosaur remains in the lower Smoky Hill Chalk
Member (Upper Coniacian) of the Niobrara Formation in western Kansas. Kansas Academy
of Science, Transactions 106(3-4):139-148.
Everhart, M. J. 2004. New data regarding the skull of Dolichorhynchops
osborni (Plesiosauroidea: Polycotylidae) from rediscovered photos of the Harvard
Museum of Comparative Zoology specimen. Paludicola 4(3):74-80.
Everhart, M. J. 2004. Plesiosaurs as the food of mosasaurs;
new data on the stomach contents of a Tylosaurus proriger (Squamata;
Mosasauridae) from the Niobrara Formation of western Kansas. The Mosasaur 7:41-46.
Everhart, M. J. 2005. Bite marks on an elasmosaur (Sauropterygia; Plesiosauria)
paddle from the Niobrara Chalk (Upper Cretaceous) as probable evidence of feeding by the
lamniform shark, Cretoxyrhina mantelli. PalArch, Vertebrate paleontology 2(2): 14-24.
Everhart, M. J. 2006. The occurrence of elasmosaurids (Reptilia: Plesiosauria)
in the Niobrara Chalk of Western Kansas. Paludicola 5(4):170-183.
Hawkins, T. 1834. Memoirs of
ichthyosauri and plesiosauri, extinct monsters of the ancient Earth. Roy. fol.,
London. ix + 58 pp., frontisp., 28 pls.
Henderson,
D.M. 2006. Floating point: a computational study of buoyancy, equilibrium, and gastroliths
in plesiosaurs. Lethaia 39:227-244.
Leidy, J. 1870. (Remarks on Elasmosaurus platyurus).
Proceedings of the Academy of Natural Sciences of Philadelphia
22:9-10. (for meeting of March 8, 1870)
Leidy, J. 1870. On the Elasmosaurus platyurus
of Cope. American Journal of Science ser. 2, 49(147):392.
Martin, J. E. and L. E. Kennedy. 1988. A plesiosaur from the late Cretaceous
(Campanian) Pierre Shale of South Dakota: A Preliminary Report. Proc. S.D. Acad. Sci.,
67:76-79
Riggs, E. S. 1939. A specimen of Elasmosaurus serpentinus. Field
Museum of Natural History, Geology 6:385-391.
Sato, T., and K. Tanabe. 1998. Cretaceous
plesiosaurs ate ammonites. Nature 394:629-630.
Schmeisser,
R.L. and Gillette, D.D. 2009. Unusual occurrence
of gastroliths in a polycotylid plesiosaur from the Upper Cretaceous Tropic Shale,
southern Utah. PALAIOS 2009 24: 453-459.
Schumacher, B. A. and M. J. Everhart. 2004. A new assessment of plesiosaurs
from the old Fort Benton Group, Central Kansas. Abstracts of oral presentations and
posters, Joint Annual Meeting of the Kansas and Missouri Academies of Science, p. 50.
Storrs, G. W. 1981. A review of occurrences of the Plesiosauria (Reptilia:
Sauropterygia) in Texas with Description of New Material. Masters Thesis, The University
of Texas at Austin, 226 pages.
Storrs, G. W. 1984. Elasmosaurus platyurus and a page from the
Cope-Marsh war. Discovery 17(2):25-27.
Storrs, G. W. 1997. Morphological and taxonomic clarification of the
genus Plesiosaurus. pp. 145-190, In Calloway, J. M. and Nicholls, E. L.,
eds, Ancient Marine Reptiles, Academic Press.
Storrs, G. W. 1999. An examination of Plesiosauria (Diapsida: Sauropterygia)
from the Niobrara Chalk (upper Cretaceous) of central North America. The University of
Kansas Paleontological Contributions, (N. S.), No. 11, 15 pp.
Taylor, M. A. 1993, Stomach stones for feeding or buoyancy? The occurrence and
function of gastroliths in marine tetrapods. Phil. Trans. R. Soc. Lond. B 341:163-175.
Taylor, M. A. 1994, Stone, bone or blubber? Buoyancy control strategies in
aquatic tetrapods, pp.151-161, In Maddock, L., Bone, Q., and Rayner, J. M. V.
(ed.), Mechanics and Physiology of Animal Swimming, Cambridge University Press.
Thompson, W. A.,
Martin, J. E. and Reguero, M. 2007. Comparison of
gastroliths within plesiosaurs (Elasmosauridae) from the Late Cretaceous marine deposits
of Vega Island, Antarctic Peninsula, and the Missouri River area, South Dakota. Pages
147-153 in Martin, J. E. and Parris D. C. (eds.), The
Geology and Paleontology of the Late Cretaceous Marine Deposits of the Dakotas. Geological
Society of America, Special Paper 427.
Welles, S. P. 1943. Elasmosaurid plesiosaurs with a description of the new
material from California and Colorado. University of California Memoirs 13:125-254.
figs.1-37., pls.12-29.
Welles, S. P. 1949, A new elasmosaur from the Eagle Ford Shale of Texas.
Fondren Science Series, Southern Methodist University, 1:1-28
Welles, S. P. 1952. A review of the North American Cretaceous elasmosaurs.
University of California Publications in Geological Sciences. 29:46-144. figs. 1-25.
Welles, S.P . and J. Bump. 1949, Alzadasaurus pembertoni, a new
elasmosaur from the Upper Cretaceous of South Dakota, Journal of Paleontology
23(5):521-535.
Williston, S. W. 1890. Structure of
the plesiosaurian skull. Science 16(407):290. (Discusses features of the skull and
turtles)
Williston, S. W. 1893. An interesting food habit of
the plesiosaurs. Kansas Academy of Science, Transactions 13:121-122, 1 plate.
(Gastroliths)
Williston, S. W. 1903. North American Plesiosaurs, Field Columbian Museum, Pub.
73, Geological Series, 2(1):1-79, 29 plates.
Williston, S. W. 1904. The stomach stones of the
plesiosaurs. Science (new series) 22:565.
Williston, S. W. 1906. North American Plesiosaurs: Elasmosaurus,
Cimoliasaurus, and Polycotylus. American Journal of Science 4(21):221-236.
Williston, S. W. 1907. The skull of Brachauchenius, with special
observations on the relationships of the plesiosaurs. United States National Museum
Proceedings 32:477-489. pls. 34-37.
Williston, S. W. 1908. North American Plesiosaurs: Trinacromerum.
Journal of Geology 16:715-735. figs. 1-15.
Williston, S. W. 1914. Water Reptiles of the Past and Present. Chicago Univ.
Press. 251 pp. (Free,
downloadable .pdf version here)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}