 |
Dr. Theophilus Turner and the
discovery of Elasmosaurus platyurus:
The rest of the story
Mike Everhart
Sternberg Museum of Natural History, Fort Hays State University, Hays, Kansas
Copyright © 2005-2017 by Mike Everhart
Created 12/09/2005: Last updated 03/06/2013
Editor's note: This web page is an updated and revised
version of: Everhart, M. J. 2005. Elasmosaurid remains from the Pierre Shale (Upper
Cretaceous) of western Kansas. Possible missing elements of the type specimen of Elasmosaurus
platyurus Cope 1868? PalArch 4(3): 19-32. A downloadable version (.pdf file,
1.2 MB) of this paper is available from PalArch, or
from the author on request. Please do not cite or quote from
this web page.
LEFT: Photograph of 27-year-old
Dr. Theophilus H. Turner (circa 1868); photo provided by Kenneth Almy. |
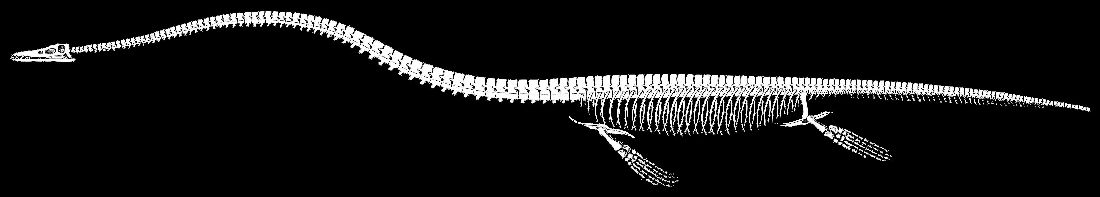 |
In 1869 when E. D. Cope described the type specimen of Elasmosaurus
platyurus Cope 1868 more completely, he noted that about four feet of dorsal
vertebrae were missing, along with the gastralia, the limbs, and most of the skull.
Although additional searches of the locality were made by Dr. Theophilus H. Turner, the
military surgeon who discovered the remains, the missing material was never located.
Interest in the specimen eventually faded as dinosaurs were discovered further West, and
portions of the specimen, including the pectoral and pelvic girdles, were mysteriously
lost. More recently, three collections of associated elasmosaurid material, including
dorsal vertebrae, ribs, gastralia, and large gastroliths were made from a second site near
the type locality of E. platyurus. The additional material is curated in the
Sternberg Museum of Natural History, Hays, KS, the University of Kansas Museum of Natural
History, Lawrence, KS, and the Cincinnati Museum Center, Cincinnati, OH. Examination the
more recently discovered remains in these three repositories, and comparisons with the
those of the type specimen at the Academy of Natural Sciences of Philadelphia, a review of
the papers, letters and other historical documents related to the discovery of E.
platyurus, and on-site evaluation of the stratigraphy of the both localities strongly
suggest that the more recently collected remains were originally part of the type specimen
and were separated prior to burial when the floating carcass began to fall apart.
Abbreviations: ANSP - Academy of Natural Sciences of Philadelphia, Philadelphia,
Pennsylvania; CMC VP - Cincinnati Museum Center, Vertebrate Paleontology, Cincinnati,
Ohio; FHSM Sternberg Museum of Natural History, Fort Hays State University, Hays, Kansas;
KUVP - University of Kansas Vertebrate Paleontology Collection, Lawrence, Kansas; NJSM -
New Jersey State Museum, Trenton, New Jersey; YPM - Yale Peabody Museum, Yale University,
New Haven, Connecticut.
INTRODUCTION
Elasmosaurus platyurus is one of the more infamous fossils associated
with the early years of paleontology in the United States, and with the "Bone
Wars" between E.D. Cope and O.C. Marsh. The story begins in western Kansas at a time
before that part of the state had been homesteaded, and before the arrival of the Union
Pacific Railroad linking Kansas City and Denver. In the spring of 1867, Dr. Theophilus H.
Turner (1841-1869) discovered the “skeleton of an extinct monster” eroding from
a ravine “some fourteen miles (22 km) north” of Fort Wallace (Almy, 1987, p.
184). In late June, he gave three of the vertebrae that he had collected to John LeConte,
a member of a party surveying the route for the Kansas (Union) Pacific Railroad (LeConte,
1868). Following the completion of the survey, LeConte delivered two (? - the fate of the
third vertebra is unknown) of the vertebrae to Edward D. Cope at the Academy of Natural Sciences of Philadelphia (ANSP) in November
of 1867. Cope (Almy 1987, p. 184) immediately recognized the vertebrae as probably
belonging to a large plesiosaur and wrote back to Turner in a letter dated December 3,
1867, asking him to procure the remainder of the specimen and send it to Philadelphia at
the expense of the ANSP. With the help of others at Fort Wallace, Turner returned to the
site of the discovery in late December, 1867, and secured some 900 pounds of bones and
concretionary matrix. Turner noted in a letter to his brother dated February 2, 1868
(ibid., p. 186) that he had collected “something over thirty-five feet of its
vertebrae”and that “there is a large amount of bony matter contained in a very hard stone matrix, some of which retains its
connection with the backbone.”
Turner was content to wait for the arrival of the railroad in late 1868. However, near the
end of February, 1868, at the urging of Cope, Turner arranged to transport the specimen by
military wagon train some 90 miles (145 km) east across the prairie to where the approaching railroad was under construction. From there, the
crates containing the remains were shipped some 1500 miles by rail to Philadelphia
(Everhart, 2005b). It appears that Cope received the specimen by mid-March, examined the
remains and made a hurried judgment as to the appearance of the animal(s) that they
represented. Then, at the March 24, 1868 meeting of the ANSP,
he reported the discovery of "an animal related to the Plesiosaurus"
which he called Elasmosaurus platyurus (Cope 1868a). A brief
note from Cope (1868b), including the new name, was also published about the same time
in LeConte's (1868) railroad survey report. In a letter to Turner dated March 25, 1868
(Almy, 1987, p. 189), Cope asked Turner "to look near where the belly of the animal
may have been and see whether there are any slender bones [gastralia], such as may occur
in the Plesiosauri." As reproduced by Almy (ibid., p. 189) Cope's note also included a sketch of what he believed
the gastralia would look like. Although Turner later returned to the site and
recovered additional material that he sent to Cope in September, 1868 (ibid., p. 193-194),
the shipment apparently did not include gastralia. Gastralia are conspicuously absent fromCope’s (1869, plate II, Fig. 1) original drawing of Elasmosaurus
platyurus and he noted (ibid., p. 44) that “the abdominal ribs [gastralia] seen
in Plesiosaurus are probably wanting as none were found by the discoverer after a
careful search.” Cope’s description of the remains in the first two versions of
his monograph (1869, p. 47 and 1870a, p. 48) indicated that he believed about 10 dorsal
vertebrae were missing in view of the fact that “Dr. Turner found that a space of
“three or four” feet intervened between the two portions of the skeleton, which
was otherwise continuous.”
The specimen (ANSP 10081) currently consists of a nearly complete vertebral column of a
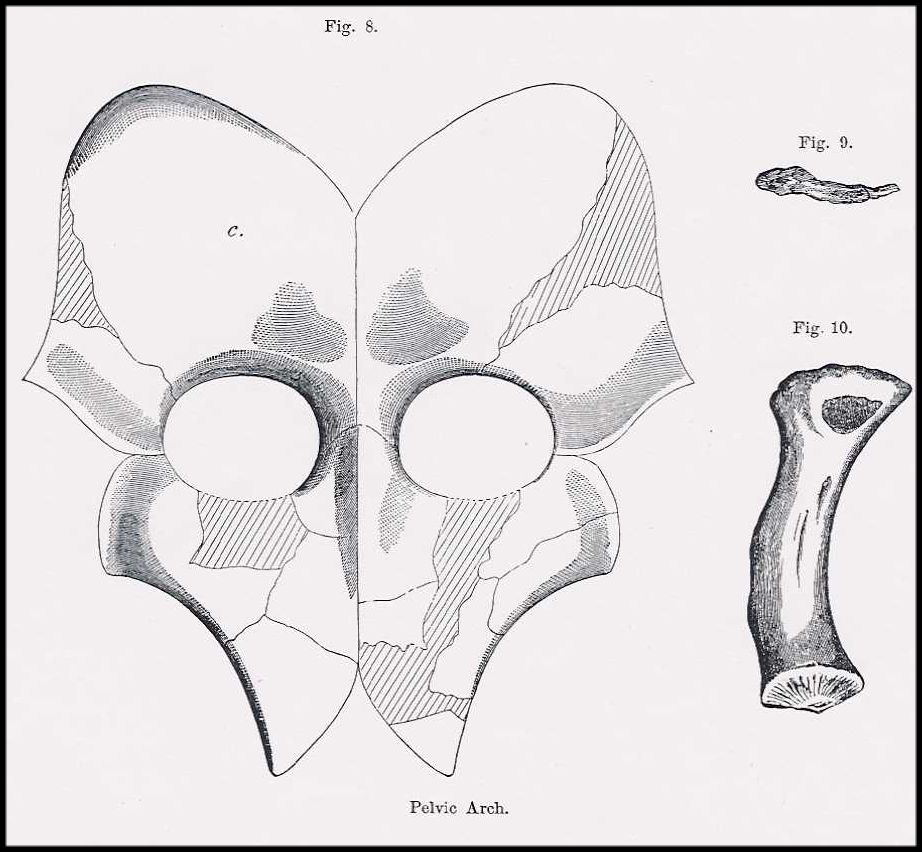
large elasmosaur, and various fragments of other bones. The pectoraland pelvic girdles, figured and described by Cope (1869 and
1870a, figure 7 and figure 8) were later lost (Williston, 1906, p. 225;
Carpenter, 1999, p. 152). Several tooth bearing fragments of the skull and lower jaws are
present, along with the occipital condyle that is still attached to the atlas-axis
vertebrae. There are only a few rib fragments remaining with the specimen and no
gastralia.
In December, 1954, George F. Sternberg received a donationof seven large (9-10 cm diameter) elasmosaurid vertebrae and "numerous" rib
fragments (FHSM VP-398) from R. B. Hooker, a landowner in Logan County, Kansas. One
vertebra had been cut lengthwise and the interior surfaces polished by Ralph Amstutz. It
is unknown if Sternberg actually visited the site, "north of McAllaster" or
conducted further investigations. One or more of
these vertebrae have been on display since that time in the Sternberg Museum of
Natural History, Hays, Kansas.
A 2002 examination of the records associated with FHSM VP-398 by the author indicated that
the locality given by Sternberg was in the vicinity of another elasmosaurid specimen
discovered in 1991 by Pete Bussen of Wallace, Kansas in a small exposure of Pierre Shale
about 1.5 km to the north and a little west of the now abandoned town site of
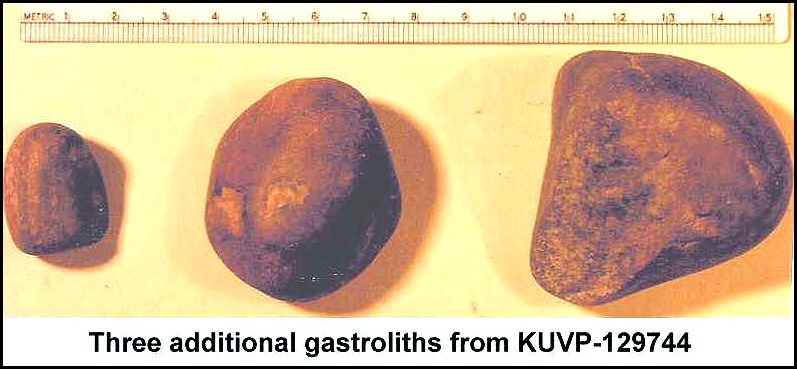
McAllaster, KS. Two dorsal vertebrae, several rib fragments and a number of unusually large gastroliths (KUVP
129744) were recovered later that year by Larry Martin of the Museum of Natural
History at the University of Kansas (Everhart, 2000). Three additional gastroliths were collected by the
author from the site in 1994. Some of these gastroliths are the largest known, and their
total weight (13.1 kg) is the heaviest ever documented from any animal (ibid.).
The dig was re-opened by Glenn Storrs and a field crew from the
Cincinnati Museum Center in 1998, and portions of eight ribs, ten gastralia and a few
probable gastroliths were recovered (CMC VP6865). Pete Bussen (pers. comm., 2002)
indicated that a local resident had also discovered a single plesiosaur vertebrae at the
same site in the 1970s, but that it had been subsequently lost.
There has been considerable erosion and excavation of the exposed Pierre Shale at the type
locality since 1868, including construction of the railroad and U. S. Highway 40, which
was originally located between McAllaster Butte and the right-of-way of the railroad. An
exploration of this locality by the author in 1999 indicated that that no visible evidence
remains of the original site where Elasmosaurus platyurus was discovered.
 |
Figure 1.
Anterior and left lateral view of the fused atlas and axis vertebrae of Elasmosaurus platyurus (ANSP 10081). The fragment
of the occipital condyle is still attached to the atlas. Note the three of the four
differing numbering systems used by various workers, including E. D. Cope on the
atlas/axis. A fourth number is on the right side. (Scale bar = 2 cm) |
 |
Figure 2. A right
oblique view of the anterior portion of the muzzle of Elasmosaurus platyurus (ANSP 10081) as collected by
Dr. Turner. Note that no complete teeth were recovered with the specimen described by
Cope, although the fresh breaks visible on the teeth suggest that they were intact at the
time of discovery. The fragment is about 11 cm in length. |
MATERIAL
ANSP 10081 – Collected by Dr. Theophilus Turner, the specimen currently consists of a
reasonably complete, articulated vertebral column (104 vertebrae, including one unnumbered and previously unreported fragment of a
cervical vertebra, but lacking many of the dorsal vertebrae), the anterior portion of
the upper and lower jaws (muzzle) that was preserved in articulation (Figure 2) and illustrated by Cope (1869, Pl. 2, figs.
8, 8a, and 8b), and four other tooth-bearing fragments
of the skull. Turner (Almy, 1987, p. 186) noted in a letter to his brother that he had
"secured part of one of its jaws containing teeth." Aside from the large jaw
fragment illustrated by Cope (1869 and 1870a, Pl. 2, Fig. 9), however, these tooth-bearing
fragments are not mentioned by Cope or later workers. The occipital condyle is also
present (Figure 1), and is still fused to the atlas-axis vertebra (Leidy, 1870, p. 9).
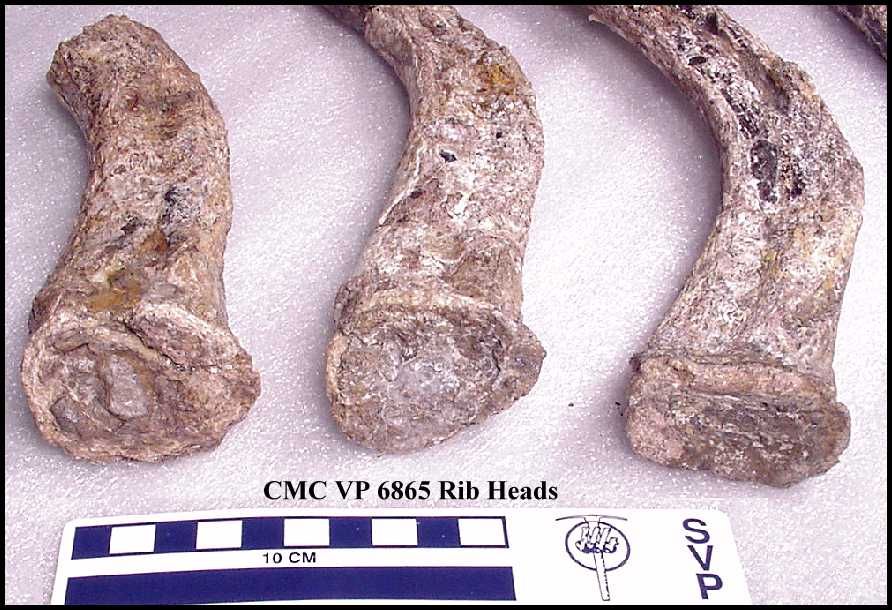
Cope (1875, p. 84) noted that the “heads of fourteen ribs are preserved, and a great
number of shafts. The heads are simple, with elongate-oval articular face.” However,
there are only a few fragments of the ribs currently
present in the specimen. The articulating surface of three remaining rib heads
measured 5.5 - 6 cm across. Many of the dorsal vertebrae and the limb girdles were
enclosed in hard, dark gray limestone concretions when collected. Some of the
concretionary material associated with the specimen
contains isolated fish teeth, scales and vertebrae that were originally described by
Cope (1868a, 1868b) and
interpreted by him as stomach contents. A single, small stone (21 x 10 x 8 mm, earlier
noted as a piece of bone by Spamer, et. al, 1995, p. 148) was found wedged in the neural canal of one of the distal caudal vertebra.
An anonymous note regarding the stone was also
included with the specimen. Based on its smooth, polished appearance, and the lack of
similar material occurring naturally in the Pierre Shale of western Kansas (Everhart,
2000), it is here interpreted to be a gastrolith.
FHSM VP-398 – Seven, large (9-12 cm diameter), mid-dorsal
vertebrae and fragments of two or more single
headed ribs collected in 1954. The articular faces of the rib heads are oval and
measure 5-6 cm across.
KUVP 129744 – Two large (9-11 cm) posterior(?)
dorsal vertebrae, fragments of single-headed
ribs, two partial gastralia and 38 gastroliths collected in 1991 by Larry Martin
(Everhart, 2000). The rib heads measure 5.5 cm.
Martin (pers. comm., 2005), recalled that both vertebrae were discovered standing on end.
CMC VP6865 – Five complete, single-headed ribs,
three partial ribs, 10 complete or nearly complete
gastralia, and seven additional gastroliths collected in 1998. The rib heads measure 5.0 – 5.5 cm across. The five
complete ribs measure 66, 66, 64, 52 and 46 cm in length. The two largest gastralia are 49
cm in length, and the smallest is about 30 cm long.
Viewed as a single specimen, the second set of remains was scattered over an approximate 4
m by 4 m area prior to burial. While several of the VP-398 vertebrae may have been
preserved together, none of the ribs or gastralia were articulated. Most of the
gastroliths, however, were located together in a single pile (Bussen, pers. comm. 1999),
suggesting that they were still contained within the plesiosaur’s digestive tract
when deposited on the sea bottom. The jumbled condition of the remains indicates that they
were most likely dropped from a floating carcass, and not scattered as the result of
scavenging. There were no bite marks on the bones and no sharks teeth recovered that would
support scavenging. While it is possible that the bones and gastroliths could have been
the last remains of a more complete individual that had mostly weathered out many years
ago, the observed taphonomy of the bones and the site do not support that scenario.
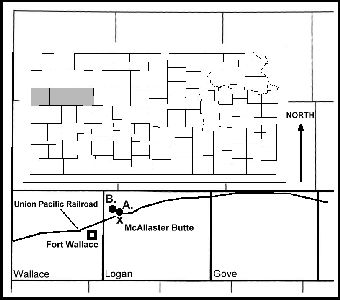 |
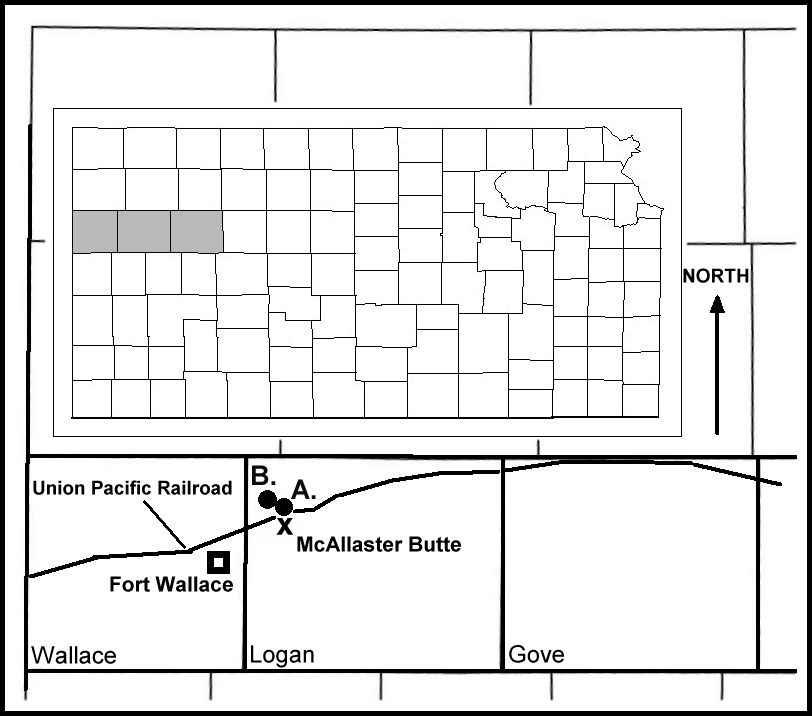
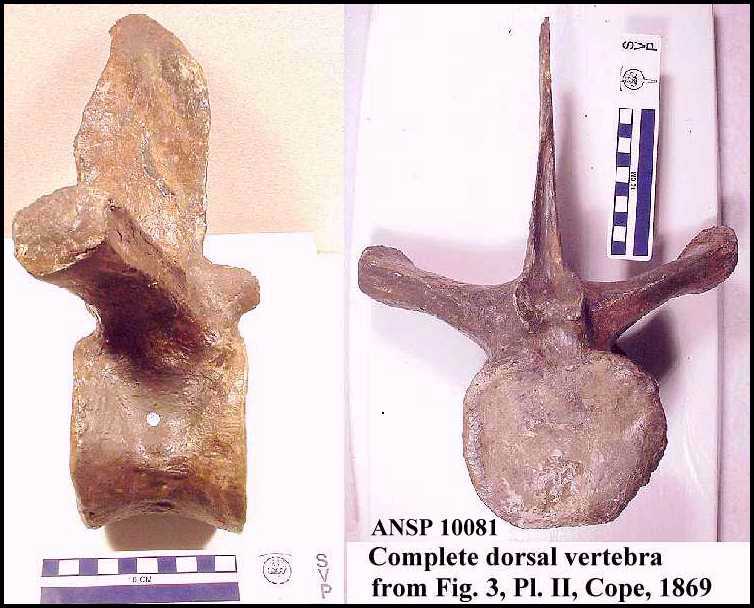
Figure 3. A map
of Kansas (inset), and Wallace, Logan and Gove counties showing the approximate locations
of: A. The type specimen of Elasmosaurus platyurus; B. Three additional sets of
elasmosaurid remains recovered since 1950; and X. McAllaster Butte. Note that the type
locality is in the opposite direction from Fort Wallace than the one recorded by LeConte
(1868). |
LOCATION / STRATIGRAPHY
From his review of the written correspondence between Turner and Cope in Almy (1987), and
his personal knowledge of the history and geography of Logan County, Pete Bussen (pers.
comm., 1998) concluded that the type specimen of Elasmosaurus platyurus had been
discovered at McAllaster Butte, a large exposure of Pierre Shale to the northeast of the
abandoned McAllaster town site (Figure 3). This is
contrary to the misleading locality information provided by LeConte (1868, p. 11) but in
keeping with the site as described in Turner's letters. On December 20, 1867, Turner
(Almy, 1987, p. 184) wrote to his brother, Daniel, about “a party of us that started
on the prairie ostensibly for the purpose of hunting but in reality for the purpose of
procuring the skeleton of an extinct monster which is embedded some fourteen miles north
of here [Fort Wallace]. It was found to rest in a slate
hill similar in appearance to those which are found on the road between home and Newton
[New Jersey].” In Turner’s letter of May 24, 1868, to his brother (Almy, 1987,
p. 191), he says that “… I think I shall do no more for the fossil till the
working party of the Rail Road reaches the spot as the present survey runs directly to the
hill and it is very probable opportunity will then offer a full exploration.” These
comments almost certainly describe the large exposure of the Sharon Springs Member of the
Pierre Shale on the north side of what is now called
McAllaster Butte in Logan County, the only notable hill along the right-of-way of the
Union Pacific railroad in western Kansas.
Based on the historical documentation, the type specimen of Elasmosaurus platyurus(ANSP 10081) was collected in 1867-68 from the north slope of what is now known as
McAllaster Butte, northeast of the town site of McAllaster, in western Logan County,
Kansas. The town of McAllaster was not established until about 1893 and has since been
abandoned. Although the exact stratigraphic occurrence of the type specimen is unknown, it
is almost certainly near the base of a large exposure of the Sharon Springs Member on the
north slope of McAllaster Butte. Turner (Almy, 1987, p. 185) noted that it “was
located in an almost perpendicular bank of [a] slate hill which made up one side of a
ravine. It was located near the bottom and required no small amount of labor in its
excavation.”
The other three specimens (FHSM VP-398; KUVP 129744 and CMC VP6865) were recovered from a
smaller shale exposure, approximately 5 m below a layer of large, septarian concretions,
and 1 m below a 2.5 cm yellow bentonite layer, about 1.5 km to the north-northwest of the
McAllaster town site. The Sharon Springs Member of the Pierre Shale (Middle Campanian) is
crops out on hillsides and along the drainage of the North Fork of the Smoky Hill River in
the immediate vicinity, and the type locality of the member is located about 3 km to the
northeast of McAllaster Butte (Elias, 1931). A town called Sheridan was established as a
camp for railroad workers on the east side of McAllaster Butte in during the construction
the Union Pacific Railroad in 1868-1869. In his fictional book called "Buffalo Land" (1872, p. 325-326), W.E. Webb
describes the discovery of a huge skeleton in a ravine ("It lay exposed, upon a bed
of slate, looking very much like a seventy-foot serpent, carved in stone") at
McAllaster Butte ("old sunken volcano"). The remains are also described as those
of a Mosasaurus, over 50 feet long, but the story easily could have been about
the type specimen of Elasmosaurus platyurus, a type of animal largely unknown
when the events in the book took place. Webb apparently had provided photographs of some
fossil bones to Leidy (1868) who briefly reported on them at
a meeting of the ANSP, but the whereabouts of the photos are unknown. A similar story was
mentioned several times by Cope in subsequent reports with providing any other details.
Cope, however,did visit the area around Sheridan in 1871 without mentioning that he was
aware that the shale near the buttes was the locality of Turner's "monster." Cope (1872, p. 325) wrote,"The yellow bed is thin or
wanting on the north bank of the Smoky, and is not observable on the north fork of that
river for twenty miles northward or to beyond Sheridan Station on the Kansas Pacific
Railroad. Two isolated hills, "The Twin Buttes," at the latter point, are
composed of the blue beds, here very shaly, to their summits. This is the general
character of the rock along and north of the railroad between this point and Fort
Wallace."
The Sharon Springs Member in Kansas was deposited in a shallow inland sea that covered
much of the central United States and Canada during middle Campanian time. According to
Parrish and Gaultier (1993), this lithostratigraphic unit is composed of soft, fissile,
bituminous shale, contains numerous bentonites and septarian concretions, and is in the
biostratigraphic zone of Baculites asperformis (Gill, et al., 1972). More
recently, however, M. Turner, et al. (2001) reported that the invertebrate fossil
assemblage in a concretion from the same stratigraphic level 3 km to the north of the type
locality was dominated by B. mclearni.
All four of the plesiosaur specimens reported here occurred below the prominent layer of
septarian concretions near the top of the Sharon Springs Member and are Middle Campanian
in age. If the second set of remains were, in fact, originally part of the type specimen,
then the current direction at the time of deposition would have been from the northwest to
the southeast. Baculites examined by Carpenter (1996) occurring in a concretion
from about the same stratigraphic level, 3 km north of the type locality, were aligned
predominantly along a northwest-southeast axis, indicating a possible direction of the
current at that time in this portion of the Western Interior Sea.
METHODOLOGY
 |
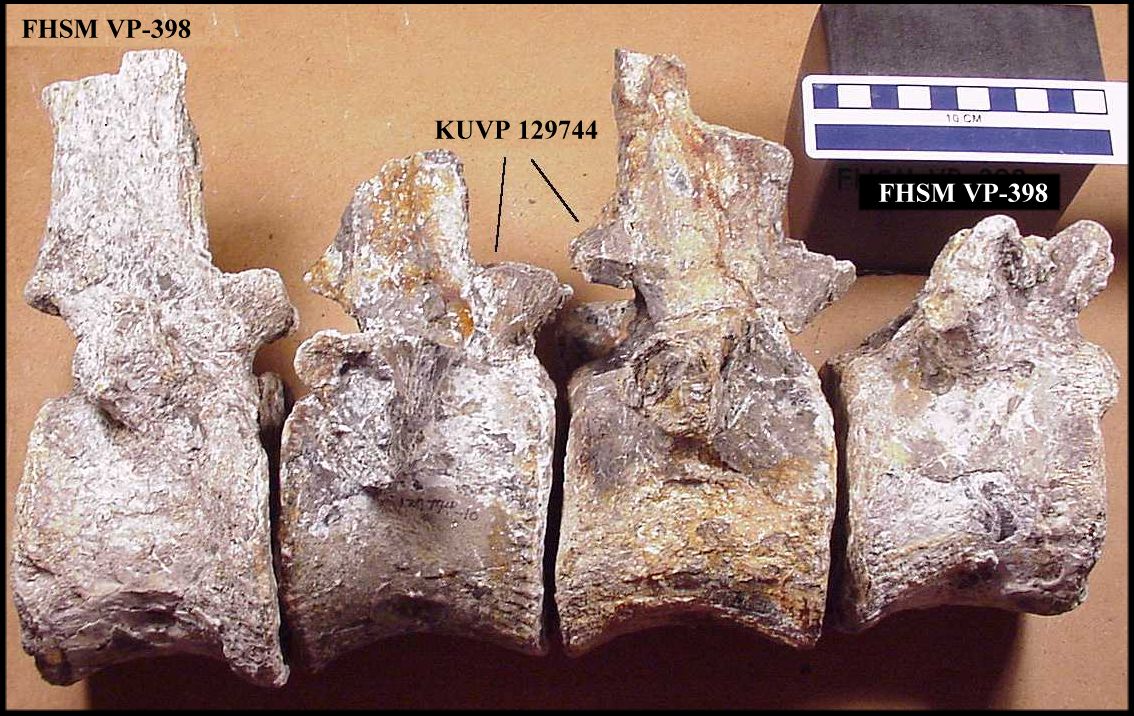
Figure 4. A. Left
lateral view of the best preserved, mid-dorsal vertebra (ANSP
10081-D3) from the type specimen of Elasmosaurus platyurus, and; B. Left
lateral view of a mid-dorsal vertebra from the second specimen (FHSM VP-398-D7). ANSP 10081-D3 was originally figured by Cope (1869, Plate 14,
fig. 3, 3a). (Scale bar = 10 cm) |
 |
Figure 5. A.
Posterior view of a dorsal vertebra from the type specimen (ANSP 10081-D4) and; B.
Posterior view of a dorsal vertebra from the second specimen (FHSM VP-398-D4). The dorsal
vertebrae of both specimens are similar in shape, size and preservation. (Scale bar = 10
cm) |
A considerable amount of historical literature (including
the 1890 exchange between Cope and Marsh in the New York Herald) is available
regarding the type specimen Elasmosaurus platyurus. However, after Leidy (1870) noted that the original description of the specimen
by Cope (1869) was reversed, Cope altered the figures and re-wrote the text of his
original pre-print (Storrs, 1984). Unfortunately, many of the details in the description
were not corrected in the subsequent revisions by Cope (1870a, 1870b, 1871), and a later
publication (1875), and cast a shadow on their usefulness. Williston (1906) provided the
next description of the type specimen but did not provide additional details or
measurements of the vertebrae. S. P. Welles(1952) re-described the specimen, revised the order and the numbering of the vertebrae,
and provided the first modern measurements of the individual vertebrae. However, he only
provided length, width and height data for one of the vertebra that he considered to be
from the dorsal series. More recently, Sachs (2005) described the remains and revised the
order of the vertebrae.
When I examined the type specimen in November, 2002, I noted that there were as many as four numbers on some vertebrae, including
Cope’s original markings. Following the most recent identification and numbering
system used by Sachs, I measured the vertebrae, directly compared them with vertebrae from
FHSM VP-398 (Figures 4 and 5), and photographed most of the specimen. I also examined bone fragments and other materials stored in a
separate container, and the remaining pieces of the gray limestone concretion that had
originally enclosed some of the bones, including one
that still contained large fragment of a vertebra.
| Table 1. - Measurements (cm) and the Roundness Index (RI = H/W)
of the dorsal vertebrae from the type specimen (ANSP 10081), and the second specimen (FHSM
VP-398 and KUVP 129744). Combined
total length = 78.7 cm / 31 in. (* =
crushed vertebra) |
| Type Specimen |
|
|
|
|
Second Specimen |
|
|
|
|
| Vertebra # |
RI |
Width |
Height |
Length |
Vertebra # |
RI |
Width |
Height |
Length |
| 10081-D1 |
0.72 |
1.25 |
9.0 |
8.7 |
VP-398-D1 |
0.91 |
10.4 |
9.5 |
8.5 |
| 10081-D2 |
0.81 |
12.0 |
9.7 |
9.1 |
VP-398-D2 |
0.78 |
11.5 |
9.0 |
8.5 |
| 10081-D3 |
0.88 |
12.0 |
10.5 |
9.1 |
VP-398-D3 |
0.79 |
12.0 |
9.5 |
9.3 |
| 10081-D4 |
0.83 |
12.0 |
10.0 |
8.7 |
VP-398-D4 |
0.82 |
11.0 |
9.0 |
8.5 |
| 10081-D5* |
0.69* |
13.0 |
9.0 |
9.2 |
VP-398-D5 |
0.92 |
11.5 |
10.0 |
9.0 |
| 10081-D6 |
0.84 |
11.9 |
10.0 |
8.6 |
VP-398-D6 |
0.91 |
11.0 |
10.0 |
9.0 |
| |
|
|
|
|
VP-398-D7 |
0.88 |
11.9 |
10.5 |
9.1 |
| |
|
|
|
|
129744-D1 |
0.89 |
10.7 |
9.5 |
8.5 |
| |
|
|
|
|
129744-D2 |
0.85 |
10.6 |
9.5 |
8.3 |
| |
|
|
|
|
|
|
|
|
|
| RANGE |
|
11.9-13.0 |
9.0-10.5 |
8.7-9.2 |
RANGE |
|
10.4-12.0 |
9.0-10.5 |
8.3-9.3 |
| AVG (n=6) |
0.80 |
12.2 |
9.7 |
8.9 |
AVG(n=9) |
0.86 |
11.2 |
9.6 |
8.7 |
The gastroliths and vertebrae collected in association with KUVP 120744 were
examined and described by Everhart (2000) and included information provided by Pete Bussen
and Larry Martin. I made on-site measurements of the CMC VP6865 specimen as member of the
1998 field crew that recovered the last portion of remains (Figure 6). In 2002, I noted
that seven plesiosaur vertebrae (FHSM VP-398) in the Sternberg Museum collection had come
from the same general locality as KUVP 129744 and CMC VP6865, and began comparing those
specimens. Subsequent visits to the University of Kansas, the Academy of Natural Sciences
of Philadelphia and the Cincinnati Museum Center provided me with the opportunity to
measure, directly compare and photograph all four of the specimens described in this
paper. Although the measurements of the dorsal vertebrae of the type specimen shown in
Table 1 are the most complete to date, the condition of those vertebrae, and that of the
two other specimens (different orientations during preservation, in or out of concretions,
etc.) convinced me that additional statistical analysis would not produce useful results.
As an example, up to 1.5 cm of the 13 cm width of vertebra 10081-D5 appears to be due to
dorso-ventral crushing.
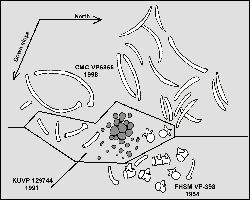 |
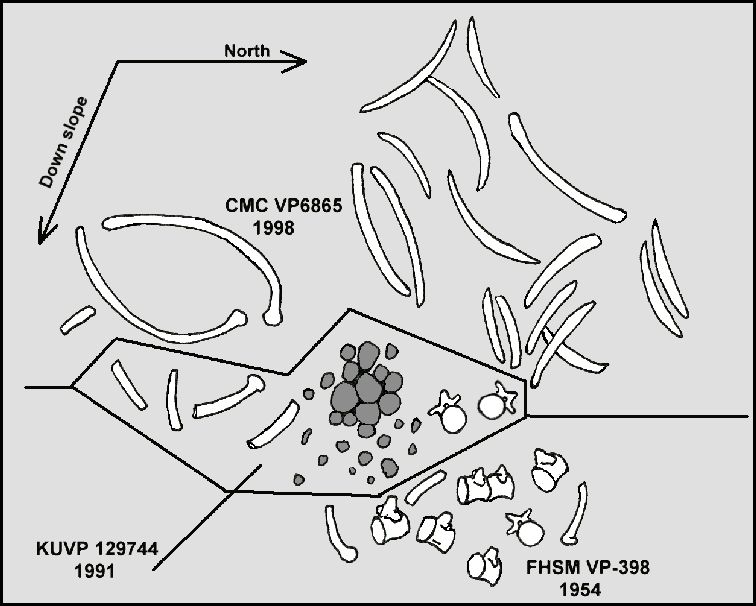
Figure 6. June 5,
1998 field sketch of the CMC VP6865 specimen by the author with the addition of the
approximate locations of the previously removed KUVP 129744 material, and conjectural
location of the seven FHSM VP-398 vertebrae. Dark gray objects near the center are
gastroliths. |
DISCUSSION
In his letter of March 25, 1868 to Dr. Turner, after his initial examination of the
remains, Cope (Almy, 1987, p. 189) wrote, "Indeed, I think that there must be a
considerable number of the vertebrae of the dorsal and cervical series remaining in the
cliff." He went on to question the orientation of the specimen and asked about the
"slender bones" [gastralia] in the stomach region. From his knowledge of English
plesiosaurs (Davidson, 2002), Cope clearly recognized that significant portions of the
original Elasmosaurus platyurus specimen were missing from the first shipment of
remains sent to him by Turner. Later, Cope (1869, p. 45-46) incorrectly concluded that the
limbs must have been small, or in the case of the hind limbs, possibly non-existent, since
he firmly believed that the animal moved by means of its long, serpent-like tail. His
infamous, head-on-the-wrong-end reconstruction (1869, pl. II,
fig. 1) does not include the rear paddles. It is likely, however, that the missing
portions, including the limbs, had simply dropped off of the bloated, floating carcass or
had been removed by scavengers before it finally came to rest on the sea bottom (Everhart,
2005a).
Carpenter (1996, p. 36) noted that in the Red Bird Member of the Pierre Shale, “the
isolated bones and skulls of vertebrates were most likely derived from floating
carcasses.” Under the right conditions, the carcass of a relatively intact elasmosaur
would likely have floated due to the decomposition gases that accumulated in the abdominal
cavity (Carpenter, 1996; Everhart, 2000, p. 72-73). It is uncertain, however, whether the
weight of the pectoral girdles, limbs and gastralia, or that of the vertebral column and
ribs would have determined the orientation (right side up or upside down) of the floating
carcass. No limb material was included in the original Elasmosaurus platyurusremains. The loss of these nearly solid, bony paddles could have affected the final
orientation of the floating carcass. In either case, the crop or stomach containing the
gastroliths would have been located just behind the pectoral girdle (Cicimurri and
Everhart, 2001). If the carcass was floating upside down, the weight of the gastroliths,
and possibly the loosened gastralia would have been resting on the dorsal vertebrae. If
the carcass was right side up, the weight of the gastroliths and the gastralia would have
been pulling down on the arch formed by the dorsal vertebrae (Taylor, 1981). This weight
would have pressed down on the abdominal wall and would have been a likely point of
failure when the integument ruptured and released the gases of decomposition. Loosened
dorsal vertebrae, gastralia and gastroliths could have then fallen out the carcass while
residual pockets of decomposition gases kept the lightened remains afloat long enough to
drift further to the southeast with the current.
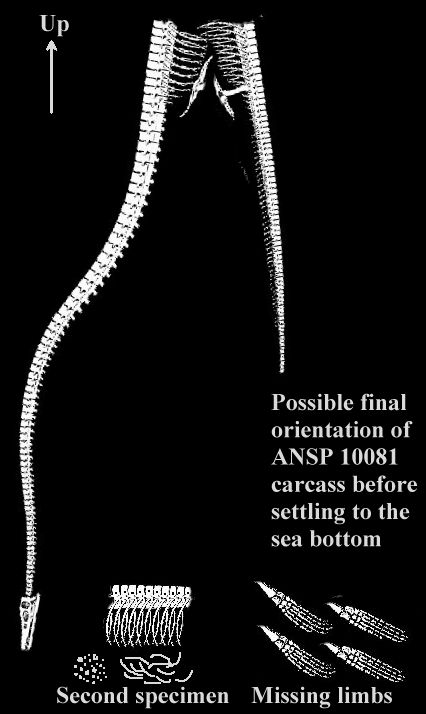
The loss of a large segment of dorsal vertebrae, ribs and gastralia from the mid-section
would have likely then caused the carcass to fold in the middle, with the long neck and head reoriented downward to a near vertical
position, roughly parallel to the caudal vertebrae. Although lightened enough to
remain suspended in the water column temporarily, the carcass would have then begun its
final descent to the bottom. The 5-6 m long neck and attached skull would have most likely
been the lowest point on the remains during the descent and would have struck the bottom
first. The impact may have been sufficient to fracture the occipital condyle and other
relatively fragile bones at the back of the elasmosaur's skull, and to detach most of the
head from the rest of the remains. The relatively "fresh" breakage apparent on
the teeth of the muzzle recovered by Turneralso suggests that they may have been taken as souvenirs by members of Turners party who
recovered the specimen in 1867-68.
While this scenario is only conjecture, it does explain many of the observations conveyed
by Turner in his letters, and the condition of the remains as reported by Cope. According
to Cope’s (1869, p. 46) original description, “Its vertebrae were found to be
almost continuous, except for a vacancy of some four feet in the anterior dorsal region.
They formed a curved line a considerable part of whose convexity was visible on the side
of a bluff of clay shale rock with seams and crystals of gypsum." Regarding the
skull, Cope (ibid.) wrote, “The end of the muzzle was broken from a part or the whole
of the cranium (Figure 2), which has not been
rediscovered, though Dr. Turner made a careful search. It was found in front of the
vertebrae here regarded as cervical, at some distance from them.” Although
Cope’s erroneous conclusion about the identification of those vertebrae (they were
caudal vertebrae, not cervical) led, in part, to his infamous head-on-the-wrong-end
reconstruction (Cope, 1869; Figure 7 herein), he failed to
modify this statement in any of the three subsequent, partially revised re-publications of
the paper (Cope, 1870a, 1870b and 1871). Since that time, the controversy that followed
regarding the initial restoration of Elasmosaurus has completely overshadowed the
fact that Dr. Turner had discovered and successfully collected one of the largest
vertebrate fossils known at the time, with no prior experience in paleontology, under
primitive conditions on the plains of western Kansas. As a comparison, the collection of a
large, nearly complete Styxosaurus snowii (NJSM 15435) from the Sharon Springs
Member of the Pierre Shale (Cicimurri and Everhart, 2001; Everhart, pers. obs.) required
several hundred hours of field time. Turner certainly deserves more recognition for this
feat than he has received to date (A brief thank you in December,
1868 from the ANSP). Dr. Turner died unexpectedly at Fort Wallace on July 27, 1869
without ever meeting Cope.
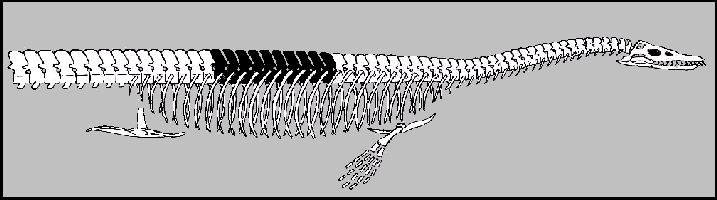 |
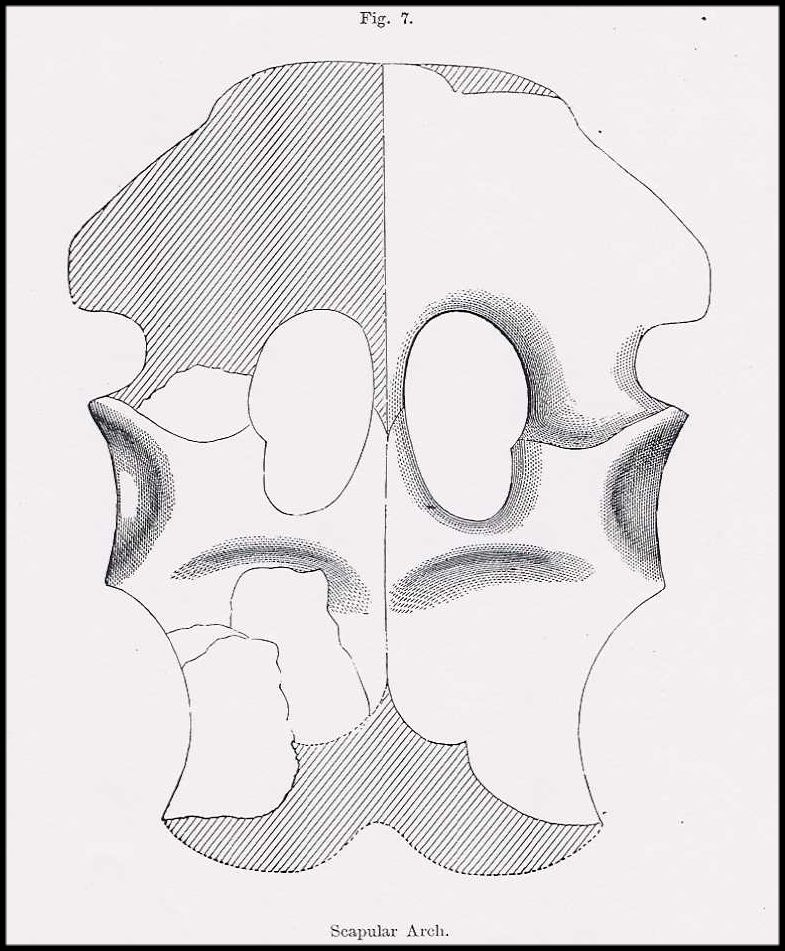
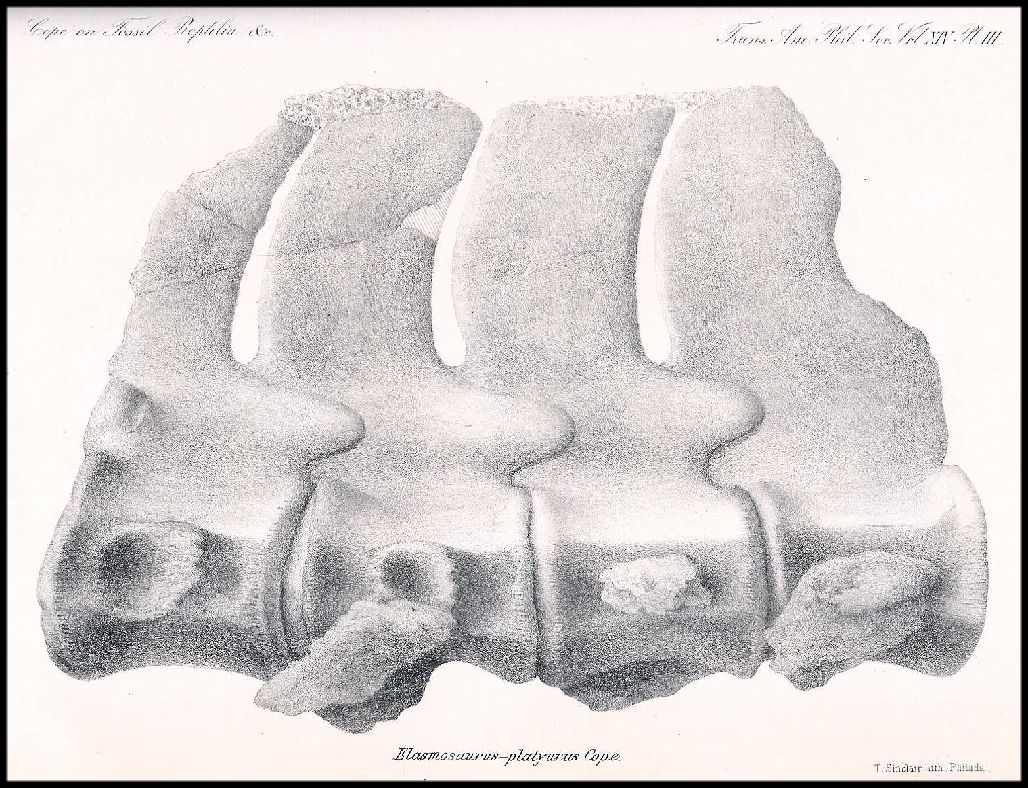
Figure 7.
Anterior portion of the "head-on-the-wrong-end" version of Elasmosaurus
platyurus published initially by Cope (1869, Pl. II, Fig. 1). The ten dorsal vertebrae
shown in black approximate those noted by Cope to be missing from the type specimen. Note
that Cope did not include the hind paddles in this figure in part due to his erroneous
belief that Elasmosaurus was propelled by its extremely long “tail”. |
Cope (1869, pl. II) figured two of the
four dorsal vertebrae that he identified without further explanation. The large vertebra
(ibid., fig. 3) shown by Cope is the most complete and least distorted of any in the
vertebrae found in the type (Figure 4A; 10081-D3,
see table 1), or the second specimen. It is apparent that some minor surface repair and
painting was done on this vertebra in Cope’s time, but its presence and excellent
condition accentuates the absence of ten or more other dorsal vertebrae from the type
specimen. Note that Welles (1952) identified five dorsal vertebrae and more recently,
Sachs (pers. comm., 2005; 2005) determined that six dorsal vertebrae were present. At
issue here is deciding where the pectoral series ends and the dorsal series begins. While
the point is arguable, the more recent examination by Sachs is accepted here.
In his description of Elasmosaurus, Cope (1869, p. 48) mentioned the “coarse
cellular texture of the spongy bone” that he had observed inside the dorsal vertebrae
where the edge of a centrum was damaged. The same
spongy bone is visible in both the ANSP 10081 vertebrae and those of FHSM VP-398. In
addition, VP-398-D5 was sectioned and polished (Figure
8) by a friend of the collector and shows that that spongy bone texture completely
fills the interior of the centrum. There is a large, medullary cavity near the center that
was apparently connected to the paired nutritive foramina that exit on the ventral side of
the vertebrae.
A comparison of the seven dorsal vertebrae of FHSM
VP-398 with the two dorsal vertebrae of KUVP 129744 shows that the centra are almost
identical shape and preservation (Table 1), with the centra of the two KU specimens being
slightly smaller. The lower elevation of the transverse processes / rib facets on the KU
specimens suggests that they are more posterior in the series than the FHSM material. Even
though they were discovered almost 40 years apart, they apparently originated from the
same locality and most probably are from the same animal. Since the KUVP 129744 and CMC
VP6865 remains were known to have come from the same site, there is no doubt that they
represent the same individual.
 |
Figure 8. Right and left halves of a dorsal
vertebra of the second specimen (FHSM VP-398-D5) as sectioned and polished soon after
their discovery. Note the spongy appearance of the bone. The lighter colored material is
calcite infilling of void spaces within the bony matrix. The large open space near the
center (medullary cavity) is connected to the two nutritive foramina that open on the
ventral surface of the vertebrae. (Scale = 10 cm) |
The composite of nine dorsal vertebrae, ribs and gastralia (hereafter referred
to as the second specimen) includes many of the same elements that Cope (1869) noted to be
missing from the type specimen of Elasmosaurus platyurus. Although Cope did not
mention gastroliths, they are now known to be routinely associated with elasmosaur remains
(Williston, 1893, 1904; Everhart, 2000, Whittle and Everhart, 2000; Cicimurri and
Everhart, 2001). With the possible exception of the small stone noted above, gastroliths
are noticeably absent from the type specimen. The unusually large size of the gastroliths
located with the second specimen is consistent with an elasmosaur of the extreme length
(45 ft; 13.7 m) suggested by Cope (1870a, p. 49). It should be noted here that Cope’s
estimate was somewhat exaggerated, and that later authors, notably Williston (1906, p.
226) and Welles, (1952, p. 54) reduced the estimated length. Based on his examination of
the original material, Welles (ibid.) also concluded that as many as 13 dorsal vertebrae
were missing from the type specimen. Measurement of all the vertebrae of the type specimen
by the author in 2001 indicated that about 9.1 m (30 ft) of the vertebral column were
present. This length, of course, does not account for the length of the skull, missing
cervical, dorsal and caudal vertebrae noted by Cope (1869) and others, or any allowances
for intervertebral cartilages between the vertebrae.
Welles (1952) provided width, height and width measurements of the vertebral centra of Elasmosaurus
platyurus (ANSP 10081) and other elasmosaurs. While few of the dorsal vertebrae of
the type specimen were included, his data for other elasmosaurs demonstrates that the
length of individual dorsal vertebrae varies by 10% or more in all of the specimens that
were measured. Recent measurements of the vertebrae of the type and second specimen (Table
1) provide an indication of both the similarity in size between the two sets of remains,
and the degree of variation in the dimensions of individual vertebra in the same animal.
Since most of the remaining dorsal vertebrae from the type specimen were apparently
discovered within a concretion while those in the second specimen were not, some
dimensional differences related to the mode of preservation would also be expected. The
average length of the vertebrae (n=9) in the second specimen was about 2% (2 mm) shorter
than the average length of the vertebrae (n=6) in the type specimen, while the average
height is only 1% (1 mm) less in the second specimen.
The greatest difference in the measurements occurs between the average widths, with those
of the second specimen being 8% (10 mm) narrower than those of the type. However,
according to Massare and Sperber (2001), elasmosaur vertebrae are normally widest over the
pectoral girdle where they would be associated with supporting the large muscles that
powered the front limbs. If the widest dorsal vertebrae were also the most anterior ones
in the type specimen, as suggested by Welles (1952), it is logical to assume that the ones
missing from the center of the vertebral column would be narrower.
The length of the centra of the six dorsal vertebrae in the type specimen differed by only
7% (6 mm) while the height differed as much as 17% (15 mm), emphasizing the variation in
the dimensions of these vertebrae in life, their different positions in the vertebral
column, the changes caused by deformation during preservation, and the small sample size.
The range of differences measured in the second specimen was similar: length – 11%
(10 mm); height – 16% (15 mm); and width – 13% (15 mm).
The average length of all the dorsal vertebrae in the type and second specimens is 88 mm.
If Welles (1952) is correct and 13 dorsal vertebrae are missing from the type specimen,
then the missing length of those vertebrae would have been about 1144 mm (45 in), not
allowing for intervertebral cartilages. If a space of 5 mm (Sato, 2003, p. 91) is included
for 13 intervertebral cartilages, the total length is increased by 65 mm (2.6 inches) to
1209 mm (47.6 in), slightly less than Cope's (1869, p. 47 and 1870a, p.48) original
estimate of 4 feet. Williston (1906, p. 226) allowed 6 mm for intervertebral cartilages
and Welles (1952, p. 54) suggested a distance of about 8 mm between the vertebrae
respectively for their estimates of the total length of the type specimen.
Measurements of plesiosaur vertebral columns by Massare and Sperber (2002) indicate that
the widest dorsal vertebrae are usually those located in the anterior part of the series,
closest to the pectoral girdle and front paddles. While Welles (1952) considered the
best-preserved vertebra of the type specimen (10081-D3) to be one of the anterior dorsals
on the basis of it being larger than the rest that he was able to measure, Sachs (pers.
comm, 2005; 2005) suggested that it is one of the mid-dorsals due to the elevated location
and greater length of the transverse processes. In view of the generally accepted idea
that the transverse processes are located at their highest point in the mid-dorsal region
of plesiosaurs, Sach’s placement of this vertebra appears to be more correct,
especially when the location of the transverse processes on 10081-D3 are compared with
those of 10081-D1. One of the vertebrae from second specimen (VP-398-D7) is nearly
identical to 10081-D3 in size and the upper location of the transverse processes, and is
here also considered to be a mid-dorsal. Damage to the dorsal processes of most of the
vertebrae in both the type specimen and the second specimen precludes a reconstruction of
the vertebral column based on the location of the transverse processes with any degree of
confidence.
The additional dorsal vertebrae (10081-D1) suggested by Sachs (pers. comm. 2005) is the
fourth in the series of four pectoral vertebraethat were preserved together and listed by Welles (1952, Table 1) as numbers 73-76 (figured by Cope). It is the first vertebra in the
series where the transverse process originates above the centrum, or about the level of
the neural canal. It is also the widest of all the vertebrae, with the exception of
10081-D5 that appears to have been crushed. Welles (1952, p. 54) also notes that the
pectoral and anterior dorsal vertebrae are the widest in the vertebral column of Elasmosaurus
platyurus, and that the width is greater than the height is greater than the length
through the dorsals, sacrals and anterior caudals. With the exception of 10081-D5, this is
the case with all of the vertebrae in the type and second specimens.
Viewed in terms of a “Roundness Index” (RI = Height/Width, where 1 = a circle),
the average “RI” of the six dorsal vertebrae in the type specimen is 0.795
(Table 1). If the crushed 10081-D5 vertebra (0.69) and the much rounder mid-dorsal
10081-D3 vertebra (0.88) are removed, the average anterior dorsal RI remains about the
same (0.80). The average RI for the second specimen (mid-dorsal and posterior dorsal
vertebrae) is 0.86, with three vertebrae scoring above 0.90. The vertebrae become rounder
through the middle of the column and then begin to flatten out again as they approach the
sacrum. This is indicted by the RI of the smallest (and probably most posterior) vertebra
(129744-D2) in the second specimen decreasing to 0.83. From the width and height
measurements provided by Welles (1952, Table 1) of four of the sacral vertebrae of the
type specimen, the RI then decreases rapidly (0.78, 0.75, 0.71, 0.67) as the vertebrae
flatten over the posterior set of paddles.
Along with the upper limb bones (propodials), the dorsal vertebrae are among the largest
and most durable bones in an elasmosaur skeleton and are some of the mostly likely bones
to be preserved due, in part, to their central location within the body. They are more
difficult for scavengers to remove than heads, tails and limbs (Everhart, 2005a). The fact
that most of the dorsal vertebrae, ribs and gastralia are missing from the type specimen
suggests that something major happened to the carcass before the remains reached the sea
bottom. While the remains of a Styxosaurus snowii specimen
(NJSM 15435) discovered by Pete Bussen about 10 km to the southwest of McAllaster
Butte were complete and generally articulated (Cicimurri and
Everhart, 2001), another Styxosaurus specimen (CMC
VP7023) recovered from the Sharon Springs Member about 3 km north of McAllaster Butte
in 1999 was more scattered (Everhart, pers. obs.). In both specimens, however, all of the
dorsal vertebrae were present and enclosed in hard limestone concretions. In contrast,
Welles (1952, p. 83) reported that the type specimen of “Elasmosaurus ischiadicus”
(YPM 1130, now Styxosaurus snowii per Carpenter, 1999) from the Smoky Hill Chalk
(Lower Campanian) is missing most of the dorsal vertebrae.
It is important to note here that the review of the material included in the type specimen
of Elasmosaurus platyurus Cope 1868 in the collection of the Academy of Natural
Sciences of Philadelphia, and that of the second specimen in the Sternberg, University of
Kansas and Cincinnati museum collections did not produce any evidence, especially
duplications of skeletal elements other than ribs, that would indicate that these were not
the remains of the same animal. Larry Martin (pers. comm., 2005) disagrees with this
premise and concluded that the catastrophic loss of this much of the skeleton from the
center of an elasmosaur would have essentially torn the animal in two and caused both
halves to sink rapidly. However, Cope’s (1869) observation that certain parts were
missing from the remains sent to him by Dr. Turner, especially the four-foot gap near the
center of the otherwise nearly complete vertebral column, suggests that the two portions
did remain attached long enough for the carcass to reach its final resting place. Although
gastralia were only included in the second specimen, the ribs, and especially the heads of
the ribs were nearly identical in size in the type and second specimens. The lithology of
the gastroliths associated with the second specimen is similar to the single stone found
wedged into the neural canal of a caudal vertebrae in the type specimen (Everhart, pers.
obs.). The similarity in size of the dorsal vertebrae, the relatively short distance
between the two localities and the apparent identical stratigraphic occurrence support the
premise that the type and the second specimen are the remains of the same elasmosaur.
CONCLUSION
While it may be impossible to prove beyond a reasonable doubt that the second specimen is,
in fact, a portion of the remains originally noted to be missing from the type specimen ofElasmosaurus platyurus Cope 1868, the evidence
strongly suggests that the two sets of remains are from the same animal. The two sites are
located within 2 km of each other and are at the same stratigraphic level. The two sets of
remains came from large elasmosaurs of approximately equal size. The type specimen was
mostly articulated when discovered while the second specimen was jumbled pile of bones and
associated gastroliths. There is no observable duplication of remains. With the notable
exception of the more readily detachable limbs, most of the bones noted by Cope to be
missing from the type specimen are present in the second specimen. Almost all of the
dorsal vertebrae (10) noted as missing from the type by Cope (1869) were recovered in the
second specimen (note that 13 vertebrae were estimated to be missing by Welles, 1952).
Since the discovery of the type specimen, gastroliths have been shown to be routinely
associated with elasmosaur remains (Williston, 1893; Everhart, 2000).
The recovery of large gastroliths in the disarticulated second specimen, and their notable
absence in the articulated remains of the type specimen begs further explanation,
especially with the recent discovery of a polished, gastrolith-like pebble in direct
association with a caudal vertebra in ANSP 10081.
Elasmosaur remains are uncommon fossils in the Western Interior Sea (see Everhart, 2006).
It would appear to be more difficult to explain the circumstances that resulted in two
sets of mutually exclusive arrays of bones from two different elasmosaurs which were
buried at approximately the same time and place in mid-ocean than to conclude that they
were from the same animal. In this case, the evidence strongly suggests that many of the
pieces missing from Dr. Turner’s original discovery of Elasmosaurus platyurushave been located.
ACKNOWLEDGEMENTS
I thank Kenneth J. Almy for publishing his genealogical research related to Dr.
Turner’s important discovery and for his generous donation of the Turner and Cope
letters in his possession to the Academy of Natural Sciences of Philadelphia; Pete Bussen
of Wallace, KS for sharing his knowledge of the history, geography and paleontology of
Logan County, and donation of the KUVP 129744 / CMC VP6865 material; Earle Spamer and Ted
Daeschler, Academy of Natural Science of Philadelphia, Philadelphia, PA for access to the
type specimen; Ken Carpenter, Denver Museum of Nature and Science, Denver, Colorado, for
the encouragement to proceed with this investigation; Larry Martin and Desui Miao,
University of Kansas Museum of Natural History, Lawrence, KS for background information
and access to KUVP 129744; Glenn Storrs, Cincinnati Museum Center, Cincinnati, OH for
discussions regarding plesiosaurs and the discovery of Elasmosaurus platyurus,
reference materials, and access to CMC VP6865; Richard Zakrzewski, Sternberg Museum of
Natural History, Fort Hays State University, Hays, KS for access to FHSM VP-398; Richard
Forest, Earth Sciences, New Walk Museum, Leicester, United Kingdom, for our discussions
regarding the various attributes of plesiosaur vertebrae; Sven Sachs, Institut für
Paläontologie, Freie Universität Berlin, Germany, for sharing data from his examination
of the type specimen; and Earl Manning, Department of Geology, Tulane University, New
Orleans, LA for reference materials and a critical review of the history of paleontology
in Kansas. Judy Massare, SUNY College at Brockport, Brockport, New York, and an anonymous
reviewer provided constructive comments that greatly improved on a previous version of
this paper.
| Editor's note: This web page is an updated
and revised version of: Everhart, M. J. 2005. Elasmosaurid remains from the Pierre Shale
(Upper Cretaceous) of western Kansas. Possible missing elements of the type specimen of Elasmosaurus
platyurus Cope 1868? PalArch 4(3): 19-32. A downloadable version (.pdf file, 1.2 MB)
of this paper is available from the author on request. Please
do not cite or quote from this web page. |
POSTSCRIPT: This paper was criticized in a following issue of PalArch (below).
The authors appear to ignore the fact that I raised the same concerns in the original
paper. There is no way of proving that the type specimen and the "second
specimen"originated from a second locality originated from the same
individual.:
Noè, L.F. & M. Gómez-Pérez. 2007. Postscript to Everhart, M.J. 2005.
“Elasmosaurid remains from the Pierre Shale (Upper Cretaceous) of western Kansas.
Possible missing elements of the type specimen of Elasmosaurus platyurus Cope
1868?” PalArch’s Journal of Vertebrate Palaeontology 4, 3: 19-32. - PalArch's
Journal of Vertebrate Palaeontology 2, 1.
The holotype is the single most important specimen in zoological taxonomy, and to avoid
confusion, it must be the remains of a single individual. Re–evaluation of data
presented to infer that three specimens collected between 1954 and 1998 are additional
material of the holotype of Elasmosaurus platyurus, indicate there is no evidence
these two sets of remains belong to the same individual, or the genus Elasmosaurus.
Historical documents indicate the missing skeletal elements of the Elasmosaurus holotype
(including dorsal vertebrae and gastralia) can be explained by factors such as weathering
and collection failure. The relative absence of gastroliths, if originally associated with
the animal, can be explained by the collecting methods employed, or the absence in
1867–1868 of a theoretical framework to explain their presence in a plesiosaur.
LITERATURE CITED:
Almy, K.J. 1987. Thof’s Dragon and the letters of Capt. Theophilus Turner, M.D., U.S.
Army. Kansas History Magazine 10(3): 170-200.
Carpenter, K. 1996. Sharon Springs Member, Pierre Shale (Lower Campanian) depositional
environment and origin of its vertebrate fauna, with a review of North American
plesiosaurs. Unpub. Ph.D. dissertation, University of Colorado, 251 pp.
Carpenter, K. 1999. Revision of North American elasmosaurs from the Cretaceous of the
western interior. Paludicola 2(2): 148-173.
Cicimurri, D.J. and M.J. Everhart. 2001. An elasmosaur with stomach
contents and gastroliths from the Pierre Shale (late Cretaceous) of Kansas. Kansas
Academy of Science, Transactions 104(3-4): 129-143.
Cope, E.D. 1868a. Remarks on a new enaliosaurian, Elasmosaurus
platyurus. Proceedings of the Academy of Natural Sciences of Philadelphia 20:
92-93.
Cope, E.D. 1868b. Note on the fossil reptiles, near Fort Wallace.
page 68, In LeConte, J. L., 1868. Notes on the geology of the survey for the extension of
the Union Pacific Railway, E. D., from the Smoky Hill River, Kansas, to the Rio Grande.
Review Printing House, Philadelphia, 76 p. with folded map.
Cope, E.D. 1869. Synopsis of the Extinct Batrachia and Reptilia of North America, Part I.
Transactions of the American Philosophical Society, New Series, 14: 1-235, 51 figs., 11
pl. (pre-print issued August, 1869)
Cope, E.D. 1870a. Synopsis of the extinct Batrachia and Reptilia of North America.
Transactions of the American Philosophical Society, New Series, 14: 1-252, 55 fig., 14 pl.
(Revised version published in April, 1870)
Cope, E.D. 1870b. Synopsis of the extinct Batrachia and Reptilia of North America.
Transactions of the American Philosophical Society, New Series, 14: 1-252, 55 fig., 14 pl.
(published in December, 1870)
Cope, E.D. 1871. Synopsis of the extinct Batrachia and Reptilia of North
America. Transactions of the American Philosophical Society, New Series, 14: 1-252, 55
fig., 14 pl.
Cope, E.D. 1875. The Vertebrata of the Cretaceous formations of the West. Report, U. S.
Geological Survey Territories (Hayden) 2: 302 pp., 57 pls.
Davidson, J.P. 2002. Bonehead mistakes: The background in scientific literature and
illustrations from Edward Drinker Cope's first restoration of Elasmosaurus platyurus.
Proceedings of the Academy of Natural Sciences of Philadelphia 152: 215-240.
Elias, M.K. 1931. The geology of Wallace County, Kansas. State Geological Survey of
Kansas, Bulletin 18, pp. 1-252, figs. 1-7, pls. 1-42.
Everhart, M.J. 2000. Gastroliths associated with plesiosaur remains
in the Sharon Springs Member of the Pierre Shale (Late Cretaceous), western Kansas.
Kansas Academy of Science, Transactions 103(1-2): 58-69.
Everhart, M.J. 2005a. Bite marks on an elasmosaur (Sauropterygia; Plesiosauria) paddle
from the Niobrara Chalk (Upper Cretaceous) as probable evidence of feeding by the
lamniform shark, Cretoxyrhina mantelli. PalArch, Vertebrate paleontology 2(2):
14-24.
Everhart, M.J. 2005b. Oceans of Kansas. A natural history of the Western Interior Sea.
Bloomington/Indianapolis, University of Indiana Press, 232 pp.
Everhart, M.J. 2005c. Elasmosaurid
remains from the Pierre Shale (Upper Cretaceous) of western Kansas. Possible missing
elements of the type specimen of Elasmosaurus platyurus Cope 1868? PalArch 4(3):
19-32
Everhart, M.J. 2006. The occurrence of elasmosaurids (Reptilia: Plesiosauria) in the
Niobrara Chalk of Western Kansas. Paludicola 5(4):170-183.
Gill, J.R., W.A. Cobban, and L.G. Schultz. 1972. Stratigraphy and composition
of the Sharon Springs Member of the Pierre Shale in Kansas. United States Geological
Survey, Professional Paper 728, 50 pp., 2 pl.
Leidy, J. 1870. [Remarks on Elasmosaurus platyurus].
Proceedings of the Academy of Natural Sciences of Philadelphia 22: 9-10.
LeConte, J.L. 1868. Notes on the geology of the survey for the extension of the Union
Pacific Railway, E. D., from the Smoky Hill River, Kansas, to the Rio Grande. Review
Printing House, Philadelphia, 76 pp. with folded map.
Massare, J.A. and S.T. Sperber. 2001. Vertebral morphology and swimming styles in
Cretaceous plesiosaurs (Reptilia, Sauropterygia). Paludicola 3(3): 95-103.
Noè,
L.F. and Gómez–Pérez, M. 2007. Postscript to Everhart, M.J. 2005. Elasmosaurid
remains from the Pierre Shale (Upper Cretaceous) of western Kansas. Possible missing
elements of the type specimen of Elasmosaurus
platyurus Cope
1868? In PalArch’s Journal of Vertebrate Palaeontology 4, 3: 19–32.PalArch’s
Journal of Vertebrate Palaeontology 2(1): 1-9
Parrish, J.T. and D.L. Gaultier. 1993. Sharon Springs Member of the Pierre
Shale: upwelling in the Western Interior Seaway? pp. 319-322 In Caldwell, W.G.E. and E.G.
Kauffman, Eds. 1993. Evolution of the Western Interior Basin. Geological Association of
Canada, Special Paper 39.
Sachs, S. 2005. Redescription of Elasmosaurus platyurus Cope 1868 (Plesiosauria:
Elasmosauridae) from the Upper Cretaceous (lower Campanian) of Kansas, U.S.A. Paludicola
5(3): 92-106.
Sachs,
S., Kear, B.P. and Everhart, M.J. 2013.
Revised vertebral count in the longest-necked vertebrate Elasmosaurus
platyurus Cope 1868, and clarification of the cervical-dorsal transition in
Plesiosauria. PLoS ONE 8(8): 6 pp. LINK
Sato, T. 2003. Terminonatator ponteixensis, a new elasmosaur
(Reptilia: Sauropterygia) from the Upper Cretaceous of Saskatchewan. Journal of Vertebrate
Paleontology 23(1): 89-103.
Spamer, E.E., E. Daeschler, and L.G. Vostreys-Shapiro. 1995. A study of fossil vertebrate
types in The Academy of Natural Sciences of Philadelphia; Taxonomic, systematic, and
historical perspectives. Academy of Natural Sciences of Philadelphia, Special Publication
16, 434 pp.
Storrs, G.W. 1984. Elasmosaurus platyurus and a page from the Cope-Marsh war.
Discovery 17(2): 25-27.
Taylor, M.A. 1981. Plesiosaurs - rigging and ballasting. Nature 290: 628-629.
Turner, M.M., K. Weed, C.D. Burke, and W. Yang. 2001. Disentangling
conflicting stratigraphic subdivisions through ammonoid biostratigraphy, the Sharon
Springs Member of the Pierre Shale Formation (Upper Cretaceous), western Kansas. GSA
2001 Rocky Mountain and South Central Sections, GSA, Joint Annual Meeting (Abstracts).
Welles, S.P. 1952. A review of the North American Cretaceous elasmosaurs. University of
California Publications in Geological Sciences 29:46-144, figs. 1-25.
Whittle, C.H. and M.J. Everhart. 2000. Apparent and implied evolutionary trends in
lithophagic vertebrates from New Mexico and elsewhere, pp. 75-82 In: Lucas, S.G. and A.B.
Heckert, Eds. 2000. Dinosaurs of New Mexico, New Mexico Museum of Natural History and
Science Bulletin 17.
Williston, S.W. 1893. An interesting habit of the plesiosaurs.
Kansas Academy of Science, Transactions 13: 121-122, 1 pl.
Williston, S.W. 1904. The stomach stones of the plesiosaurs.
Science (New Series) 22: 565.
Williston, S.W. 1906. North American plesiosaurs: Elasmosaurus, Cimlioasaurus,
and Polycotylus. American Journal of Science, Series 4, 21(123): 221-234, 4 pl.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}