
|
Copyright © 2004-2014 by Mike Everhart
Page created 09/26/2004 / Last updated 01/11/2014
 |
LEFT: In the National
Geographic IMAX movie Sea Monsters (2007), there is a Tylosaurus
lurking somewhere under the surface, waiting for its next meal including a
small polycotylid plesiosaur (Dolichorhynchops osborni) shown
overhead. While not entirely correct scientifically regarding the behavior
of this mosasaur, the movie itself is based on the actual 1918 discovery
of a Tylosaurus proriger skeleton with the bones of a juvenile
plesiosaur inside the rib cage. The discovery was made by Charles H.
Sternberg in Logan County, Kansas, and the specimen was sold to the United
States National Museum (USNM - Smithsonian) where it has been on display
since about 1920.
Although Sternberg published a short note about the discovery, it was more or less lost in history until I came across the report in the Transactions of the Kansas Academy of Science (1922): I joined them in the Kansas chalk in Logan county, on Butte creek. I was so fortunate as to find a fine tylosaur skeleton the second day in the field. There were twenty-one feet of the skeleton present in fine chalk. The complete skull was crushed laterally. Nearly the complete front arches and limbs were present, as was also the pelvic bones and both femora. All the vertebrae to well into the caudal region beyond the lateral spines were continuous, with the ribs in the dorsal region. Between the ribs was a large part of a huge plesiosaur with many half-digested bones, including the large humeri part of the coracoscapula, phalanges, vertebrae, and, strangest of all, the stomach stones, showing that this huge tylosaur, that was about twenty-nine feet long, had swallowed this plesiosaur in large enough chunks to include the stomach. How powerful the gastric juice that could dissolve these big bones! This specimen I sent to the United States National Museum. |
Please do not cite or quote from this page. This webpage is an updated version of:
Everhart, M. J. 2004. Plesiosaurs as the food of mosasaurs; new data on the stomach contents of a Tylosaurus proriger (Squamata; Mosasauridae) from the Niobrara Formation of western Kansas. The Mosasaur 7:41-46.
Copies of the printed article are available as a .pdf file (Adobe Reader) on request.
Abstract Although plesiosaurs and mosasaurs co-existed for about 25 million years at the end of the Late Cretaceous, the fossil record was mute regarding interactions between these two groups of marine reptiles until a discovery in the summer of 1918. At that time, Charles F. Sternberg uncovered the partially digested bones of a plesiosaur as stomach contents in an adult (9 m) Tylosaurus proriger skeleton in the Smoky Hill Chalk Member (Early Campanian) of the Niobrara Formation near Twin Butte Creek in Logan County, Kansas. Sternberg reported his discovery at the annual meeting of the Kansas Academy of Science in 1919 and indicated that the material had been sent to the United States National Museum. Due to unusual circumstances regarding the publication of his brief paper in the Transactions of the Kansas Academy of Science, however, the association of the two specimens went largely unnoticed until 2001. This unusual association demonstrates conclusively that Tylosaurus fed on plesiosaurs and provides additional data about the ecology of the Western Interior Sea. Here the remains are re-examined and discussed in light of related information that has become available in the more than eighty years since their original discovery. |
Introduction
The remains of plesiosaurs (Sauropterygia) are found in the fossil record from the Late Triassic through the end of the Cretaceous. They evolved into many diverse forms, including the giant pliosaurids, Kronosaurus and Brachauchenius, polycotylids (Trinacromerum and Dolichorhynchops) and the extremely long-necked elasmosaurids. Plesiosaurs, in general, survived several near extinctions, the latest occurring in the Early Cretaceous (Bakker, 1993). During the last 25 million years of the Cretaceous, the two kinds of plesiosaurs remaining were the ‘long-necked,’ small-headed elasmosaurids and the ‘short-necked’, long-headed polycotylids. In the Late Cretaceous Western Interior Sea, elasmosaurids such as Elasmosaurus platyurus Cope 1868 and Thalassomedon haningtoni Welles 1943 reached lengths of 14 m (45 ft.), whereas the much smaller polycotylids such as Dolichorhynchops osborni Williston 1902 grew only as large as 5-6 m (16-20 ft.). Both groups fed predominantly on small, nektonic fish and cephalopods (Brown, 1904; Massare, 1987; Cicimurri and Everhart, 2001), and would have been relatively harmless to larger fish and other marine reptiles.
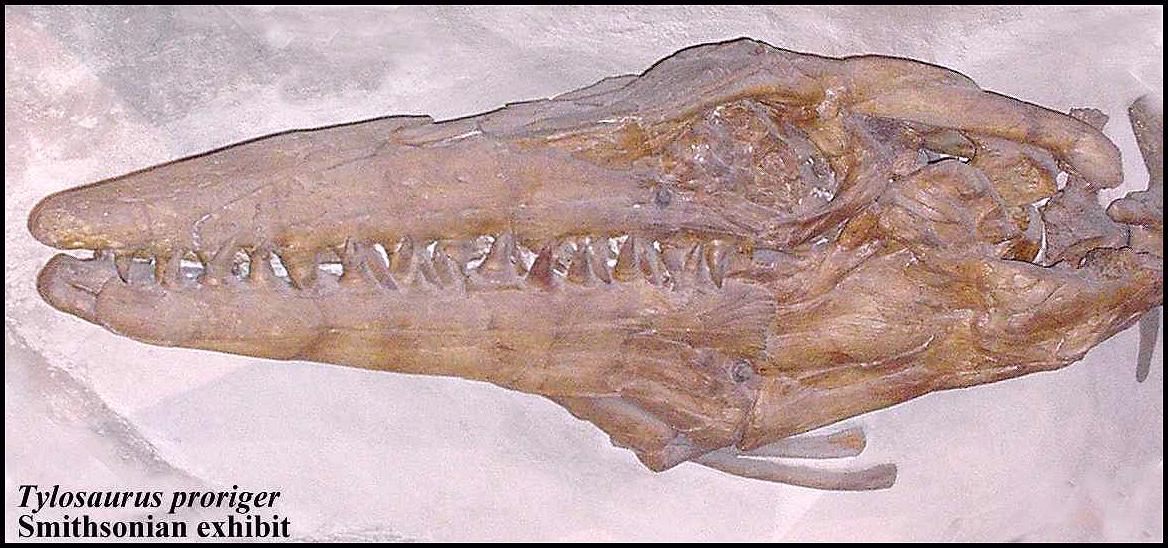
Mosasaurs were the top predators in the oceans of the Late Cretaceous, with some species growing to lengths of 17 m / 55 ft. (Lingham-Soliar, 1999) during the Maastrichtian, and were well adapted for eating large prey. Most mosasaurs during the Early Campanian, such as Platecarpus and Clidastes, were were somewhat smaller and were not known to have fed on anything other than fish. A highly flexible, kinetic skull and additional joint in the lower jaw (Russell, 1967), however, allowed mosasaurs to swallow large prey whole, very much like the feeding adaptations in snakes. Williston (1898, p. 213) suggested that while "a saurian of the largest size [Tylosaurus] might swallow something as large as a two-year-old calf," he believed that a mosasaur's "food was evidently the numerous small fishes that swarmed the seas, with perhaps an occasional animal of their own kind." More recently, Lee, Bell and Caldwell (1999, p. 656) proposed that the "highly flexible lower jaw" evolved in mosasaurs "for accommodating large prey."
From preserved gut contents associated with mosasaur remains, these marine reptiles are known to have fed on a variety of prey, including fish, marine birds, sea turtles, and other mosasaurs. Williston (1898) and Camp (1942) reported fish scales and bones within specimens of Tylosaurus and Plotosaurus, respectively. Williston (1899, p. 40-41) described the stomach contents of a well-preserved Platecarpus mosasaur that contained the remains of 1.2 m (4 ft) fish (Cimolichthys), and Everhart (unreported data) found partially digested Cimolichthys vertebrae in association with the skeleton of a large (9 m) Tylosaurus proriger (FFHM 97-10). The partially digested remains of a teleost fish (Bananogmius), a marine bird (Hesperornis), a small mosasaur (Clidastes) and a shark (teeth of Cretalamna?) were reported by Martin and Bjork (1987) as the preserved stomach contents of a large Tylosaurus proriger from the Pierre Shale (Middle Campanian) of South Dakota. Dollo (1887) identified turtle remains associated with a specimen of the giant tylosaurine, Hainosaurus bernardi, from Upper Maastrichtian deposits in Belgium. Bite marks attributable to a giant Maastrichtian mosasaur, such as Prognathodon or Mosasaurus, are visible on a carapace fragment of large marine turtle (Allopleuron hoffmanni, #7431) in the Teylers Museum, Haarlem, The Netherlands (pers. observation, 2004). From the evidence in the fossil record, it is readily apparent that mosasaurs, especially the larger individuals, were capable of swallowing large prey whole. Varricchio (2001, p. 405) indicated that "extant archosaurs, crocodilians and birds, typically ingest prey with little mastication and can fully digest bones." It is likely that mosasaurs were also capable of completely digesting the bones of large prey.
 |
In 1918, Charles H. Sternberg and his sons (George F. and Levi)
recovered a nearly complete skeleton of a large, adult Tylosaurus
proriger that included the partially digested bones of a plesiosaur (Sternberg, 1922). Sternberg reported the unique specimen at the
51st annual meeting of the Kansas
Academy of Science in 1919, and stated that the specimen had been sent to the United
States National Museum. His brief note, however, was not published until 1922 (Volume 30,
including the years 1919-1921) because of administrative problems between the Kansas
Academy of Science and the State Printer (Skelton, 1998). From
the lack of recognition of this discovery in more recent literature, it is evident that
the single paragraph in the brief, two-page paper drew little attention outside that of
the membership attending the meeting in 1919. The mosasaur and plesiosaur remains were received by the National Museum in 1919 and curated separately. The mosasaur (USNM 8898) became a permanent display in the Ancient Seas exhibit at the museum where it is today, whereas the plesiosaur bones (USNM 9468) were stored in the collections. Gilmore (1921) reported on the preparation of the Tylosaurus specimen, and the food habits of mosasaurs, without mentioning the association with the plesiosaur. The USNM records for both specimens, however, do specifically record their association. Sternberg’s hand-written field tag is included with the plesiosaur remains. It reads; "Half digested plesiosaur bones between the ribs of No. 17. Also pebbles from the stomach of plesiosaur. C. H. Sternberg." Although the Tylosaurus proriger skeleton was listed by Russell (1967, p. 174), its connection with the plesiosaur was not mentioned. In any case, predation or scavenging by mosasaurs on plesiosaurs is not documented in recent publications. |
Abbreviations
AMNH – American Museum of Natural History, New York, NY; FHSM – Fort Hays State University Sternberg Museum of Natural History, Hays, KS; KUVP – University of Kansas Museum of Natural History, Lawrence, KS; LACMNH – Los Angeles County Museum of Natural History, Los Angeles, CA; MCZ – Museum of Comparative Zoology, Harvard University, Cambridge, MA; USNM – United States National Museum, Washington, D. C.
Locality and Stratigraphy
The USNM records state that the remains were discovered on the south side of [Twin] Butte Creek, 15 miles southeast of Russell Springs, in Logan County, Kansas. The locality data, and the chalky matrix remaining on the plesiosaur bones, confirm that the specimen came from the Smoky Hill Chalk Member of the Niobrara Chalk Formation. Both Hattin (1982) and Bennett (2000) consider this general area to be in the upper one-third of the chalk, probably in the vicinity of Hattin’s marker unit 15 or 16. The locality suggests that the age of this specimen is probably late Santonian or early Campanian. Stewart (1990, p. 25) noted that the "nearly all the partial skeletons of Niobrara polycotylids have been collected from the upper horizons of the Smoky Hill Chalk Member." According to Storrs (1999), both elasmosaurid and polycotylid plesiosaurs have been reported from the upper Smoky Hill Chalk Member; however, the available records indicate that polycotylid remains are the more common. The type specimen of Polycotylus latipinnis Cope 1869 (AMNH 1735/USNM 244534) was discovered in Logan County by William. E. Webb (Cope, 1870, p. 34-38, pl. 1, figs. 1-12). The type of Dolichorhynchops osborni Williston 1902 (KUVP-1300) at the University of Kansas Museum of Natural History, the mounted specimen of D. osborni (FHSM VP-404) at the Sternberg Museum of Natural History, and the juvenile D. osborni remains (MCZ-1064) at the Harvard Museum of Comparative Zoology were also from the upper chalk of Logan County. Thirty years after the first discovery of a plesiosaur in Kansas, Williston (1897) interpreted the upper chalk as being deposited in shallow water nearer to shore than the lower chalk.
Although plesiosaur remains are very rare in the lower chalk, Stewart (pers. comm., 2002) has collected a juvenile propodial from Sand Creek in southern Rooks County near the contact of the Smoky Hill Chalk Member with the Fort Hays Limestone (late Coniacian). In 1990, I observed J. D. Stewart collect an sub-adult Dolichorhynchops propodial and associated epipodials (LACMNH 148920) in the early Santonian chalk of Lane County. Everhart (2003) reported 5 sets of fragmentary plesiosaur remains (FHSM VP-13962 to 13966), including fragments of a Dolichorhynchops skull and lower jaws, from the late Coniacian and early Santonian lower chalk of Gove County .
The first Cretaceous reptile reported from Kansas, Elasmosaurus platyurus Cope 1868, was collected from the Sharon Springs Member of the Pierre Shale Formation northeast of Fort Wallace, Kansas by Dr. Theophilus H. Turner (Cope, 1868) in 1867-1868. Although thousands of mosasaur remains have been recovered from the Smoky Hill Chalk Member (Everhart, 2001) since Tylosaurus proriger Cope (MCZ-4374) was described in 1869, only a few dozen plesiosaur specimens have been discovered there. The rarity of plesiosaurs suggests that they were only occasional inhabitants of the central portion of the Western Interior Sea over what is now Kansas, and possibly were only migrating across that large body of open water to reach feeding grounds closer to either the eastern and western shores.
Discussion
Sternberg (1922) reported the discovery of the large, well-preserved Tylosaurus proriger specimen in an chalk exposure along [Twin] Butte Creek in Logan County, Kansas. The 29 ft. (8.8 m) skeleton included an articulated skull, front limbs, ribs, pelvic bones and both femora, and all but the last 6 ft (1.9 m) of the tail. In between the ribs was "a large part of a huge plesiosaur with many half digested bones, including the large humeri, part of the coracoscapula, phalanges, vertebrae, and, strangest of all, the stomach stones, showing that this huge tylosaur … had swallowed this plesiosaur in large enough chunks to include the stomach (Sternberg, 1922, p.120)." Sternberg, however, did not comment as to whether he believed the plesiosaur had been live prey or a scavenged carcass.
 |
In November, 2001, I examined the plesiosaur specimen (USNM 9468).
Although the bones show extensive surface erosion and represent only a small portion of
the entire skeleton, they appear to be those of a polycotylid, Dolichorhynchops osborni.
The remains are consistent with the description provided by Sternberg (1922), except that
it is not a "huge" individual. Based on measurements of the two, reasonably
complete humeri, the plesiosaur was somewhat smaller than either the type specimen
(KUVP-1300) or the mounted specimen in the Sternberg Museum (FHSM VP-401). All of the
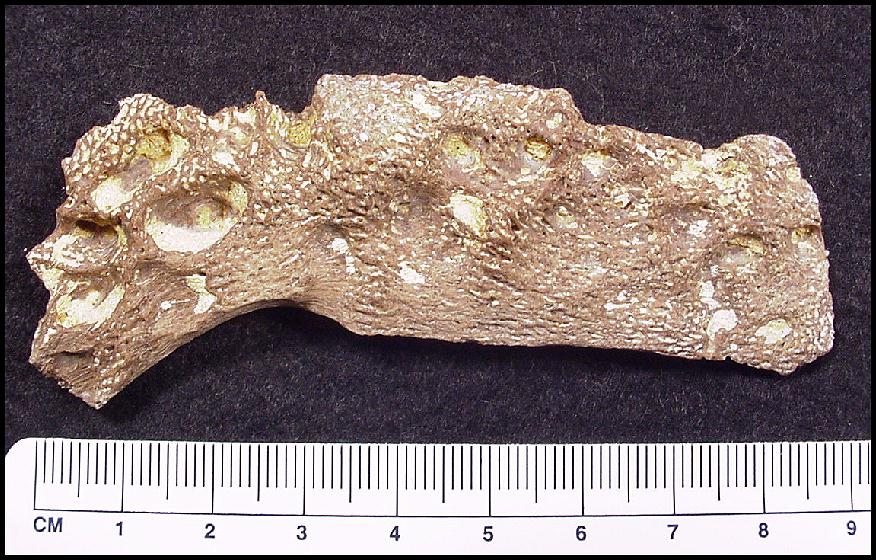
bones show evidence of some dissolution of the surface layer by stomach acids (Fig. 1). LEFT: Figure 1 - Unidentified plesiosaur bone fragment showing loss of surface layer and deep pitting due to attack by stomach acids. (Scale = mm) |
The damage is similar to the surface erosion described on hadrosaur bones identified as gut contents of a tyrannosaurid (Varricchio, 2001, p. 401-402). He noted that sub-surface bone structures such as the "vascular canals and intertrabecular spaces have been enlarged, giving the bone a spongy appearance clearly distinct from fresh bone." None of the plesiosaur bones appear to have been weathered and /or sun-bleached (as opposed to being corroded by stomach acids) and from Sternberg’s description (1922, p.119), it is likely that the plesiosaur’s bones were still completely embedded in the chalk matrix when discovered.
Some of the plesiosaur bones appear to be heavily eroded on one side and relatively undamaged on the other, a possible indication that they were partially protected from attack by stomach acids by attached muscle, other undigested tissue or adjacent remains. Although no tooth marks are evident on the bones, the presence of numerous small fragments argues that some of the larger bones in the pectoral or pelvic girdles were crushed or broken when the plesiosaur was bitten as it was being swallowed by the mosasaur. Bite marks that were present originally may also have been removed by the digestive process. In addition to corrosion by stomach acids, portions of the relatively thin right coracoid appear to have been damaged during recovery and / or storage. Many bone fragments are sufficiently corroded as to make identification difficult, including some relatively thick pieces that are probably parts of the missing femora.
 |
The more cartilaginous ends of the humeri are severely eroded
(Fig. 2a; Fig. 2b), whereas the dense perichondral bone on the shafts appears to have been
more resistant to the stomach acid (O'Keefe, pers. comm., 2002). They measure 26 and 27 cm
in length and, taking into account the damage to both ends, were probably no longer than
30 cm in life. LEFT: Figure 2a -Dorsal and ventral views of the left humerus, showing extensive damage to the cartilaginous ends of the bone and relatively little erosion of the denser perichondral bone of the shaft. (Scale = cm) RIGHT: Figure 2b - Dorsal and ventral views of the right humerus. |
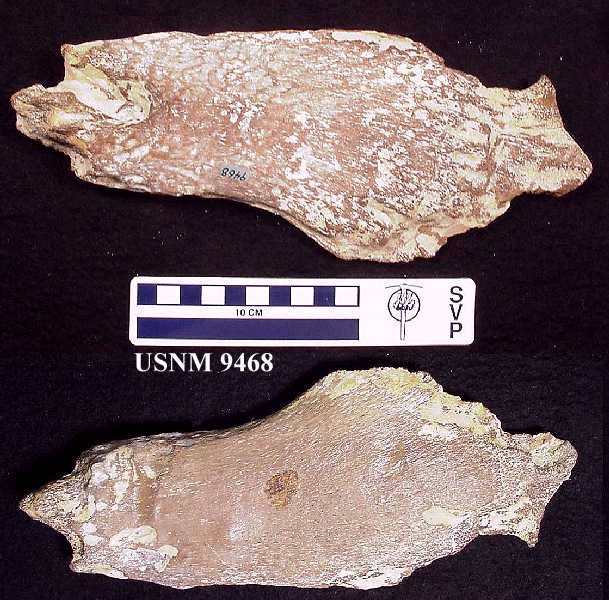 |
This compares favorably with the length of the right humerus (33.6 cm) of the 3 m specimen of Dolichorhynchops osborni (FHSM VP-404; Bonner, 1964, p. 53) in the Sternberg Museum collection, indicating that the plesiosaur was somewhat less than 3 m (9 ft.) in length. Three mesopodials / metapodials are also included in the limb elements. Twenty-seven podials (phalanges) were also recovered from the remains, far fewer than the number (70+) normally present in a single paddle of Dolichorhynchops.
 |
There are only four, small caudal (?) vertebrae (22-27 mm
diameter) associated with the specimen and all appeared to have been badly corroded by
acid. The small size of the vertebrae and the
presence of several, small, caudal ribs (Fig. 3) indicates that they were from near
the end of the tail. However, Bruce Schumacher (pers. comm. 2004) thinks that these may
instead be cervical vertebrae on the basis on the large nutritive foramina. The damage,
potentially including enlargement of the nutritive foramina, caused by the digestive processes of the mosasaur make positive identification
difficult. LEFT: Figure 3. A ventral view of four partially digested caudal vertebrae and three caudal ribs. A small gastrolith (arrow) remains attached to one of the ribs. (Scale = mm) |
 |
Sternberg (1922, p. 120) reported that stomach
stones (gastroliths) were collected with the specimen (Fig. 4). Although there was no
notation of gastroliths indicated in the USNM records, 12 small (up to 28 mm in length),
mostly siliceous stones were included with the remains along with two small concretionary
masses that appeared to contain smaller grains of quartz sand.
LEFT: Figure 4. Small gastroliths and two larger masses (left) containing sand grains recovered from the plesiosaur remains. (Scale = mm) |
Quartz or quartzite sand and gravel is not a normal component of the limestone or chalk beds that make up the Niobrara Formation (Mudge, 1877; Hattin, 1982) and the nearest sources would have been located hundreds of miles away (Williston, 1893). With the likely exception of a group of 12 rounded and polished stones (FHSM VP- 13919), from the lower Smoky Hill Chalk Member (late Coniacian) in Gove County, the only other gastroliths (a shark - KUVP 68979; and an elasmosaur - YPM 1644) known from the Niobrara Chalk were associated with vertebrate remains. As noted by Sternberg (1922), the presence of gastroliths suggests that the plesiosaur’s gut was intact when it was swallowed by the Tylosaurus.
The largest USNM 9468 gastrolith measures 28 x 14 x 8 mm. Two of the smaller gastroliths (8-9 mm maximum length) were still embedded in the chalk matrix and closely associated with partially digested bone fragments. Another small stone is pressed tightly against a caudal rib. In the late Cretaceous Western Interior Sea, gastroliths are most often associated with the remains of long-necked elasmosaurid plesiosaurs, such as Styxosaurus and Thalassomedon (Cicimurri and Everhart, 2001), and are only rarely documented from the short-necked, polycotylid group (see Martin and Kennedy, 1988). Although 12 gastroliths is a small number compared to the hundreds that have been discovered with the remains of other plesiosaurs (Everhart, 2000), they have never been reported from a mosasaur. It is most likely that the gastroliths were from the plesiosaur and not from the larger mosasaur that ate it.
A single, un-eroded Squalicorax falcatus tooth is also included with the plesiosaur remains and was likely shed during post-mortem scavenging of the Tylosaurus carcass by these sharks. Squalicorax bite marks have often been observed on vertebrate remains in the Smoky Hill Chalk Formation (Schwimmer, Stewart and Williams, 1997). Scavenging of the unprotected and readily removable viscera of the mosasaur by these sharks (Cicimurri and Everhart, 2001) could also explain why more of the plesiosaur remains were not recovered with the Tylosaurus skeleton.
In addition, half of a large (antero-posterior length = 47 mm; diameter = 66 mm) and partially digested teleost vertebra (Xiphactinus audax) was also included with the plesiosaur specimen (USNM 9468). Although the corroded appearance of the vertebra is similar to the condition of the plesiosaur bones, it is also weathered, and may not have been collected with the specimen. It was not mentioned in Sternberg's (1922) report.
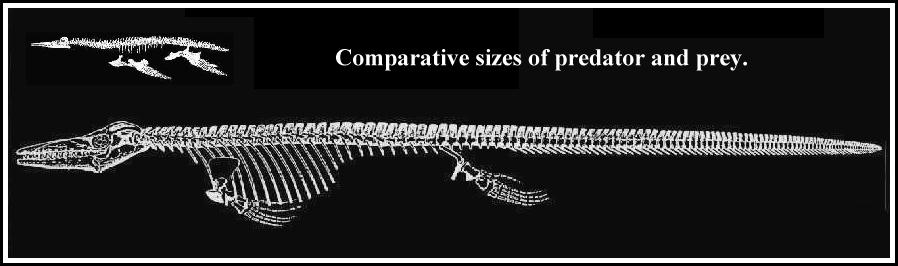
A method which measures the amount of water displaced by a scale model (Alexander 1989) was used to estimate the weights of the mosasaur and the plesiosaur. This approach assumes that the density of the animal is approximately the same as water (1 gm/cc). A 9 m Tylosaurus proriger would have weighed about 1100 kg while a 2.5 m pliosaur would have weighed between 60-70 kg. While this would have been a very large meal for the mosasaur to swallow, the evidence appears to support that it did occur.
The acid-etched condition of the plesiosaur remains, whether it was killed by the mosasaur or scavenged, suggests that the plesiosaur had been consumed several hours prior to the death of the Tylosaurus. There is no indication of what may have killed the mosasaur and postmortem scavenging by sharks appears to be have been limited to the viscera, the missing rear paddles and the distal caudal elements. The relative completeness of the mosasaur skeleton as described by Sternberg (1922) suggests that the remains reached the sea bottom shortly after death and it is likely that intact ribcage of the much larger mosasaur may have prevented this relatively small portion of the plesiosaur’s remains from being carried away by the scavengers.
Conclusion
Partially digested plesiosaur bones were discovered within the rib cage of an articulated, adult Tylosaurus proriger by Charles Sternberg in 1918 and reported by him at the 1919 meeting of the Kansas Academy of Science. His brief paper describing the remains was published in 1922 and remained largely unnoticed since that time. This specimen provides the first evidence that the larger individuals of Tylosaurus at least occasionally fed on marine reptiles besides turtles and smaller mosasaurs. That a 9 m mosasaur (skull length = 1.2 m) was able to swallow all, or a large portion of, a 3 m plesiosaur in mid-ocean, provides additional data as to the predatory / scavenging habits and feeding capabilities of these large marine animals. Although predation or scavenging by mosasaurs on plesiosaurs should not be unexpected, the rarity of preservation of stomach contents in the fossil record and an unusual combination of modern circumstances have delayed prior recognition of Charles H. Sternberg’s important discovery.
Acknowledgements
I am obliged to Robert Purdy and Michael Brett-Surman, (United States National Museum, Washington, D.C.), for their assistance in locating and providing data on the USNM collection and access to this specimen. Larry Martin and Desui Miao, (University of Kansas, Lawrence, KS), provided access to relevant specimens in the collection in the Museum of Natural History. F. Robin O’Keefe, (New York College of Osteopathic Medicine, Old Westbury, NY) shared age, condition and identification information from his earlier examination of the specimen. In addition to the data provided by J. D. Stewart, (Los Angeles County Museum of Natural History, Los Angeles, CA), about Niobrara plesiosaurs, I have greatly benefited from our many hours of discussions in regard to the biostratigraphy of the Smoky Hill Chalk Member. I also thank Earl Manning (Tulane University, New Orleans) and Richard Zakrzewski (Sternberg Museum, Fort Hays State University) for their reviews and comments on the original manuscript.
Literature Cited / Suggested Reading:
Alexander, R. M. 1989. Dynamics of dinosaurs and other extinct giants. Columbia University Press, New York, 167 pp.
Bakker, R. T. 1993. Plesiosaur extinction cycles - Events that mark the beginning, middle and end of the Cretaceous. pp. 641-664, In Caldwell, W. G. E. and E. G. Kaufmann, (eds.), Evolution of the Western Interior Basin, Geological Association of Canada, Special Paper 39.
Bennett, S.C. 2000. Inferring stratigraphic position of fossil vertebrates from the Niobrara Chalk of western Kansas. Kansas Geological Survey, Current Research in Earth Sciences, Bulletin 244, part 1.
Bonner, O. W. 1964. An osteological study of Nyctosaurus and Trinacromerum with a description of a new species of Nyctosaurus, Unpublished Masters Thesis, Fort Hays State University, 63 pp.
Brown, B. 1904. Stomach stones and the food of plesiosaurs. Science, 20(501):184-185.
Camp, C. L. 1942. California mosasaurs, University of California Press, Berkeley and Los Angeles, CA, 67 pp.
Cicimurri, D. J. and Everhart, M.J. 2001. An elasmosaur with stomach contents and gastroliths from the Pierre Shale (Late Cretaceous) of Kansas. Transactions of the Kansas Academy of Science 104(3-4):129-143.
Cope, E. D. 1868. [Remarks on a new large enaliosaur]. Proceedings of the Academy of Natural Sciences of Philadelphia 20:92-93.
Cope, E. D. 1870. Extinct Batrachia, Reptilia and Aves of North America. Transactions of the American Philosophical Society. N. S. 14:1-253.
Dollo, L. 1887. Le Hainosaure et les nouveaux vertébrés fossiles du Musée de Bruxelles. Revue des Questions Scientifiques XXI 504-539; XXII, 70-112. (in French; Hainosaurus and new vertebrate fossils of the Museum of Brussels)
Everhart, M. J. 2000. Gastroliths associated with plesiosaur remains in the Sharon Springs Member of the Pierre Shale (late Cretaceous), western Kansas. Kansas Academy of Science, Transactions 103(1-2):58-69.
Everhart, M. J. 2001. Revisions to the biostratigraphy of the Mosasauridae (Squamata) in the Smoky Hill Chalk Member of the Niobrara Chalk (late Cretaceous) of Kansas. Kansas Academy of Science, Transactions 104(1-2):56-75.
Everhart, M. J. 2003. First records of plesiosaur remains in the lower Smoky Hill Chalk Member (Upper Coniacian) of the Niobrara Formation in western Kansas. Kansas Academy of Science, Transactions 106(3-4):139-148.
Everhart, M. J. 2004. Notice of the
transfer of the holotype specimen of Niobrarasaurus coleii (Ankylosauria;
Nodosauridae) to the Sternberg Museum of Natural History. Kansas Academy of Science,
Transactions 107(3-4):173-174.
Everhart, M. J. 2004. Conchoidal fractures preserved on elasmosaur gastroliths are
evidence of use in processing food. Journal of Vertebrate Paleontology 24 (Supplement to
3):56A.
Everhart, M. J. 2005. Bite marks on an elasmosaur (Sauropterygia; Plesiosauria) paddle
from the Niobrara Chalk (Upper Cretaceous) as probable evidence of feeding by the
lamniform shark, Cretoxyrhina mantelli. PalArch, Vertebrate paleontology 2(2):14-24.
Everhart, M. J. 2005. Rapid evolution, diversification and distribution of mosasaurs
(Reptilia; Squamata) prior to the K-T Boundary. Tate 2005 11th Annual Symposium in
Paleontology and Geology, Casper, WY, pp. 16-27 (not peer-reviewed).
Everhart, M. J. 2005. Oceans of Kansas - A Natural History of the Western Interior Sea.
Indiana University Press, 322 pp.
Gilmore, C.W. 1921. An extinct sea lizard from western Kansas. Scientific American CXXIV 273, 280, 3 figs.
Hattin, D. E. 1982. Stratigraphy and depositional environment of Smoky Hill Chalk Member, Niobrara Chalk (Upper Cretaceous) of the type area, western Kansas. Kansas Geological Survey Bulletin 225, 108 pp.
Lee, M.S., Bell, G.L., Jr. and Caldwell, M.W. 1999. The origin of snake feeding. Nature, 400:655-659
Lingham-Soliar, T. 1999. What Happened 65
Million Years Ago: The Study of Giant Marine Reptiles Throws New Light on the last Major
Mass
Extinction, Science Spectra, No. 17:20-29.
Martin, J. E. and Bjork, P.R. 1987. Gastric residues associated with a mosasaur from the late Cretaceous (Campanian) Pierre Shale in South Dakota. In: Martin, J. E. and G. E. Ostrander, editors, Papers in Paleontology in Honor of Morton Green, Dakoterra 3. pp. 68-72.
Martin, J. E. and Kennedy, L.E. 1988. A plesiosaur with stomach contents from the late Cretaceous (Campanian) Pierre Shale of South Dakota: A preliminary report. Proceedings of the South Dakota Academy of Science 67:76-79.
Massare, J. A. 1987. Tooth morphology and prey preference of Mesozoic marine reptiles. Journal of Vertebrate Paleontology 7(2):121-137.
Mudge, B.F. 1877. Notes on the Tertiary and Cretaceous periods of Kansas. In: Part I (Geology) of the Ninth Annual Report, U. S. Geological and Geographical Survey of the Territories (Hayden), for 1875, pp. 277-294.
Russell, D.A. 1967. Systematics and morphology of American mosasaurs. Peabody Museum of Natural History, Yale University, Bulletin 23, 241 pp.
Schwimmer, D.R., Stewart, J.D. and Williams, G.D. 1997. Scavenging by sharks of the genus Squalicorax in the Late Cretaceous of North America, Palaios, 12:71-83.
Skelton, L. H. 1998. A brief history of the Kansas Academy of Science. Kansas Academy of Science, Transactions 101(3-4):140-145.
Sternberg, C.H. 1922. Explorations of the Permian of Texas and the chalk of Kansas, 1918. Kansas Academy of Science, Transactions 30(1):119-120.
Stewart, J.D. 1990. Niobrara Formation vertebrate stratigraphy. In: Bennett, S. C., editor, Niobrara Chalk Excursion Guidebook, University of Kansas Museum of Natural History and Kansas Geological Survey, pp 19-30.
Storrs, G. W. 1999. An examination of Plesiosauria (Diapsida: Sauropterygia) from the Niobrara Chalk (upper Cretaceous) of central North America. University of Kansas Paleontological Contributions, New Series, 11:15 pp.
Varricchio, D. J. 2001. Gut contents from a Cretaceous tyrannosaurid; implications for theropod dinosaur digestive tracts. Journal of Paleontology, 75(2):401-406.
Williston, S.W. 1893. An interesting food habit of the plesiosaurs. Kansas Academy of Science, Transactions 13:121-122, 1 pl.
Williston, S.W. 1897. The Kansas Niobrara Cretaceous, The University Geological Survey of Kansas, 2:237-246.
Williston, S.W. 1898. Mosasaurs. In: Williston, S. W. (ed.), The University Geological Survey of Kansas, 4:81-347, pls. 10-72.
Williston, S.W. 1899. Some additional characters of the mosasaurs, The Kansas University Quarterly, 8(1):39-41, 1 plate.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}