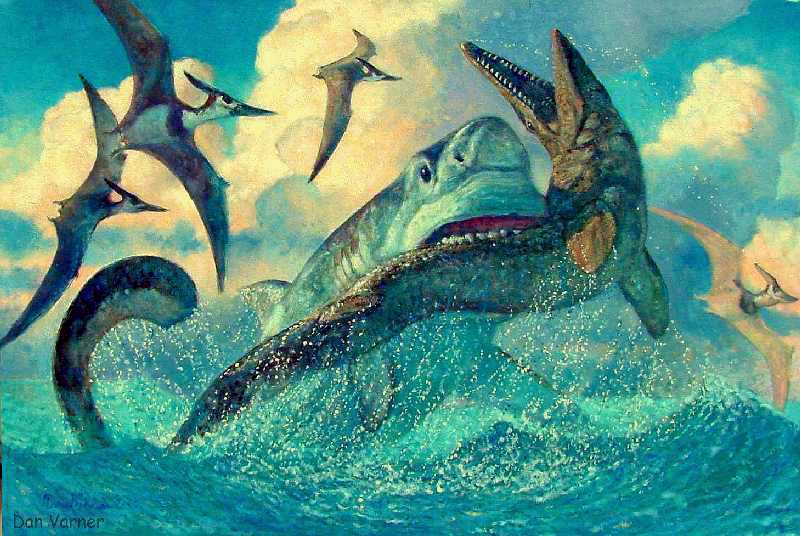 |
Mosasaurs: Last of the great
marine reptiles
Copyright © 2000-2009 by Mike Everhart
Last updated: 07/18/2009
Original article published as: Everhart, M. J. 2000. Mosasaurs:
Last of the great marine reptiles. Prehistoric Times. 44:29-31
Copyright © 1999 by Dan Varner; used
with permission of Dan Varner |
I was pleased to see mosasaurs being featured in the June/July 2000 (#42)
edition of Prehistoric Times and wanted to take the opportunity to update the article with
some additional information. As webmaster at Oceans of Kansas Paleontology, and an Adjunct
Curator of Vertebrate Paleontology at the Sternberg Museum of Natural History, my
interests are centered on late Cretaceous marine life in general and mosasaurs in
particular. Mosasaurs were the dominant predator in the oceans of the late Cretaceous and
were certainly the marine equivalent of Tyrannosaurus rex. The
finding of the first mosasaur remains in the Netherlands about 1780 (Mosasaurus
hoffmanni) predates the discovery of dinosaurs by more than fifty years, yet
they have been frequently ignored in the media and text books on paleontology in favor of
the more popular ichthyosaurs and plesiosaurs. Maybe it is because they look superficially
like alligators and crocodiles, or it might be that they are simply too common, since they
are represented in the fossil record from every continent by thousands of specimens from
more than 40 known species. In spite of this lack of good press, however, there have been
a number of recent, exciting discoveries that add to our knowledge of these magnificent
animals.
UPDATE 2005: This article was extensively rewritten and cited, and now is
available as Chapter 9 of my book Oceans of Kansas - A
Natural History of the Western Interior Sea
See also: Everhart, M. J. 2005. Rapid evolution,
diversification and distribution of mosasaurs (Reptilia; Squamata) prior to the K-T
Boundary. Tate 2005 11th Annual Symposium in Paleontology and Geology, Casper, WY, p.
16-27 (not peer-reviewed).
The ancestors of
mosasaurs were probably related to small terrestrial lizards called aigialosaurs that
lived close to the ocean from the late Jurassic through the Cretaceous. In the
mid-Cretaceous, the ancestral mosasaurs began to rapidly adapt to life in the ocean. They
were not the first reptiles to leave the land and return to the sea, but they were
possibly the most successful. Ichthyosaurs and plesiosaurs had inhabited the oceans since
the Triassic, evolving into many diverse forms and surviving several major extinction
events. For unknown reasons, ichthyosaurs declined significantly in early Cretaceous and
are thought to have been extinct about the time that the earliest mosasaurs re-entered the
water. Plesiosaurs were also less numerous in the late Cretaceous than during the
Jurassic, and had evolved into some very specialized forms like the long-necked elasmosaurs. Even the short-necked plesiosaurs (pliosaurs and polycotylids) were much smaller than their Jurassic
cousin, Liopleurodon, and an early Cretaceous
relative, Kronosaurus. It is possible that both the
ichthyosaurs and the plesiosaurs were losing the evolutionary
battle of "who eats who" to faster, larger and more advanced varieties of fish
such as Xiphactinus and the giant
Ginsu sharks (Cretoxyrhina mantelli). Several other groups of reptiles,
including marine crocodiles, teleosaurs, placodonts and turtles had also enjoyed limited successes in the marine
environment, but none approached the world-wide domination that mosasaurs would attain in
the late Cretaceous.
The early ancestors of mosasaurs probably fed in the
ocean and returned to land much like the marine iguanas that are found today in the
Galapagos Islands. Over a relatively short period of time, however, these ancestral
mosasaurs became larger and more specialized, evolving rapidly into several genera of
highly successful predators. By the beginning of Coniacian time (about 90 million years
ago - mya), there were three major genera (Tylosaurus, Platecarpus and Clidastes)
living in the Western Interior Seaway. Tylosaurs were the largest of these early
mosasaurs, reaching about 10 meters (30 feet) in length by Santonian time (85 mya). Platecarpus
was probably the most common of the first wave of mosasaurs and reached lengths of about 8
meters (24 feet). Clidastes, the smallest and probably the most primitive of the
group, was generally less than 5 meters in length when full grown (specimens of larger
individuals are known). By the Campanian, Tylosaurs were even larger (13-14 meters) and
many more species were making their appearance. Within the space of a few more million
years, by Maastrichtian time (70 mya), mosasaurs were truly huge, with several lineages (Mosasaurus
and Hainosaurus – a close relative of Tylosaurus) reaching 15 meters
(50 feet) or more. One giant specimen (Hainosaurus bernardi) found in Europe was 17
meters (almost 55 feet) in length. There was no doubt who were the biggest and baddest
predators in the oceans 70 million years ago.
As mosasaurs increased in size and diversity during
the last 25 million years of the Cretaceous, they also spread around the world. While
their fossilized remains have been found most often in Kansas and South Dakota, their
bones are known from every continent, and were most recently found in Antarctica. It is
interesting to note that the word "mosasaur" means Meuse River lizard, a tribute
to the locality where the first mosasaur was found in the
Netherlands. (Read a translation of the original story here)
Soon afterwards, mosasaur remains were discovered in England and as far away as New Zealand. The first North American mosasaur remains (mostly
teeth) were described from the greensands of New Jersey. In the early 1830s, a German
nobleman, Prince Alexander Philip Maximilian von Wied and a Swiss artist, Karl Bodmer,
traveled among the Indians in wilds of Nebraska and South Dakota. Along the way, they
acquired the remains of a Mosasaurus that had been found
along the Missouri River in what would become South Dakota. That specimen eventually
found its way back to Germany where it was prepared and described in an 1845 paper by Dr.
August Goldfuss. The skull of this North American Mosasaurus is still in the
collection of the Goldfuss Museum in Bonn, Germany. In 1869, E. D.
Cope began briefly described and named the first mosasaur from Kansas and the rush for
fossils was on. His rival, O. C. Marsh, organized the Yale College Scientific Expeditions
of the 1870s and collected about two thousand mosasaur specimens from Kansas for the
Peabody Museum.
Although they probably preferred the shallower coastal
waters where their prey was most abundant, they were also excellent open water swimmers.
Mosasaurs were certainly capable of traveling across large bodies of water as evidenced by
the numerous fossils found in the chalks and shales deposited in the middle of the Western
Interior Seaway, hundreds of miles from the nearest land. From the number of remains that
are found in these deposits, it seems apparent that mosasaurs were living in mid-ocean,
and not just migrating through it. Fossils found recently in Canada and in Africa suggest
that near the end of the Cretaceous, some species were adapting to freshwater habitats
that had been the primary domain of alligators and crocodiles.
Mosasaurs used a side to side, undulating motion of
their long, sinuous tails to swim through the water. They were probably not capable of
swimming long distances at high rates of speed but rather hunted from ‘ambush’,
surprising and out-accelerating or outrunning their prey over a short distance. The tail
of Clidastes is somewhat shorter in proportion to its body length than that of
other mosasaurs, but has elongated vertebral processes that would have supported a much
deeper tail. This may indicate that this genera was faster than other mosasaur genera.
Speed may have been a very useful adaptation when your larger cousins (tylosaurs) saw you
as their next meal. However, it was not always enough as demonstrated by fossilized stomach contents of a tylosaur found in
South Dakota which included a smaller Clidastes.
Because mosasaurs were so well adapted to living in
the ocean, it is improbable that they ever voluntarily came out on land. As they adapted
to life in the ocean, their legs and feet became broad, flat paddles used primarily for
steering. Their limbs were no longer solidly connected to their backbones and could not
support the weight of their large bodies out of the water. While they may have been able
to slither through shallow water much like a snake, they would have been nearly helpless
on dry land. Once beached, they would have most likely died of suffocation from the
collapse of their lungs, or from overheating.
While their bones show that mosasaurs were certainly
reptiles, the discovery of a Plioplatecarpus
‘mother mosasaur’ in South Dakota with the remains of several unborn young in
her abdomen appears to show conclusively that they bore their young live. Like
ichthyosaurs, mosasaurs probably only gave birth to a maximum of 4 or 5 babies at time.
This is a very small number compared to other reptiles such as crocodiles, alligators and
turtles that lay dozens of small eggs, and depend on sheer numbers for the survival of
their young. Early paleontologists, such as Samuel Williston, concluded that young
mosasaurs must have lived in estuaries or other near-shore, protected environments because
their remains had not been found along with those of the adult mosasaurs. Whether those
remains were simply overlooked or ignored by Cope, Marsh, Sternberg and others in the rush
for bigger, more spectacular specimens, we know now that mosasaurs of all ages lived and
died in the middle of the Western Interior Seaway.
The presence of the remains of young individuals in
the Smoky Hill chalk of Kansas indicates that mosasaurs were giving birth in mid-ocean,
200 miles or more from the nearest land. It also suggests that mosasaurs, out of
necessity, may have lived together in groups for the protection of their young. There were
too many other hungry predators around, including 20 foot long sharks, giant predatory
fish (Xiphactinus) and other species of mosasaurs, for
young mosasaurs to have survived long without some assistance. While it is possible that
the Western Interior Seaway had masses of floating seaweed that young animals could hide
in, there is no fossil evidence to support this theory and the presence of many fast
swimming predators, such as Xiphactinus, seems to argue for large areas of open,
unobstructed water. One variety of primitive swordfish (Protosphyraena)
had long pectoral fins that extended two to three feet outward on either side of its body,
hardly a good design for efficient hunting in a kelp forest. Most other marine reptiles,
including turtles, polycotylids and plesiosaurs, also moved through the water with long,
outstretched limbs.
Mosasaurs probably fed primarily on fish, but are
known to have eaten just about anything that they could swallow. The preserved stomach
contents of a large tylosaur on exhibit in the Museum of Geology at
the South Dakota School of Mines and Technology contains the bones of a smaller
mosasaur (Clidastes), a bird (Hesperornis), a
fish (Bananogmius) and possibly a shark. Fish remains were found in a specimen of Plotosaurus
from California and a Tylosaurus from Kansas. The remains of a turtle are preserved
in an European specimen, and the exhibit specimen of Tylosaurus
proriger at the Smithsonian was discovered with a plesiosaur as its last meal.
Ammonite shells have been found with what appear to be mosasaur bite marks, but there is
some question as to whether or not these holes were caused by mosasaur bites or far more
likely, by the attachment of limpets. A Globidens specimen
found in South Dakota has the ground of remains of clams in its abdomen.
It is important to note here that, with the exception
of a few specialized and late evolving species, mosasaur jaws and teeth were used for
seizing and holding of prey until it could be swallowed. Most mosasaur teeth are cone
shaped and do not have the cutting edges typical of shark or theropod teeth. Short of
dismembering its prey by the sheer force of its bite, a mosasaur had to swallow whatever
it caught whole, much like a modern snake. Mosasaurs and snakes are also similar in the
way that their skulls are constructed, including a lower jaw with a flexible joint that
was used to ‘ratchet’ prey into the mosasaur’s throat with the aid of an
additional, double row of teeth in the roof of their mouth. Living in mid-ocean, such
adaptations were necessary for survival, and mosasaurs were winners in that regard. Only a
few of the later genera of mosasaurs developed specialized teeth for cutting
or tearing flesh (Leiodon), or for crushing clams and other hard shelled
invertebrates (Globidens).
| "They were furnished, like snakes, with
four rows of formidable teeth on the roof of the mouth. Though these were not designed for
mastication, and, without paws for grasping, could have been little used for cutting, as
weapons for seizing their prey they were very formidable. And here we have to consider a
peculiarity of these creatures, in which they are unique among animals. Swallowing their
prey entire like snakes, they were without that wonderful expandability of throat due in
the latter to an arrangement of levers supporting the lower jaw. Instead of this each half
of that jaw was articulated or jointed at a point nearly midway between the ear and the
chin. This was of the ball-and-socket type, and enabled the jaw to make an angle outward,
and so widen by much the space enclosed between it and its fellow. The arrangement maybe
easily imitated by directing the arms forward, with the elbows turned outward and the
hands placed near together. The ends of these bones were in the Pythonomorpha as
independent as in the serpents, being only bound by flexible ligaments. By turning the
elbows outward and bending them, the space between the arms becomes diamond-shaped and
represents exactly the expansion seen in these reptiles, to permit the passage of a large
fish or other body. The arms, too, will represent the size of jaws attained by some of the
smaller species. The outward movement of the basal half of the jaw necessarily twists in
.the same direction the column-like bone to which it is suspended. The peculiar shape of
the joint by which the last bone is attached to the skull depends on the degree of twist
to be permitted, and therefore to the degree of expansion of which the jaws were
capable." E. D. Cope, 1872 |
Life in the oceans of the Cretaceous was dangerous at
best and mosasaurs did not always have things their way. New evidence, including sharks
teeth embedded in partially digested mosasaur bones, from the late Coniacian Smoky Hill
Chalk of Kansas indicates that mosasaurs were frequently fed upon by the large ginsu
sharks (Cretoxyrhina mantelli). While it is not possible
at this point to determine whether this feeding activity was the result of an attack on a
live mosasaur or the scavenging of a carcass, it is probable that a five or six meter
Ginsu would behave much like a modern Great White shark, or an extinct Carcharodon
megalodon. More than likely, Ginsu sharks attacked small, injured or sickly mosasaurs
when they most vulnerable, but probably would not have passed up a free meal either. The
rest of the story, however, indicates that Cretoxyrhina became extinct over the
space of a few million years at the same time that mosasaurs were becoming bigger and much
more wide spread. Did mosasaurs cause the extinction of these large sharks? No one knows
for sure, but we do know that modern sharks suffer greatly from over-fishing because they
do not reproduce rapidly. It seems reasonable that bite-sized, juvenile sharks were on the
menu for growing numbers of large, hungry mosasaurs.
Mosasaurs ruled the oceans of the Late Cretaceous and were beginning to invade
fresh water environments such as estuaries, swamps and rivers when the Age of Dinosaurs
ended. Did they die suddenly due the catastrophic effects of an asteroid impact in the
Yucatan, or was their extinction more gradual following the general collapse of the marine
ecosystem? We may never know.
Go here for
a webpage that describes the meteor impact that caused the Manson Crater in Iowa, 74
million years ago
Acknowledgments: Although not mentioned in the published article, I would like
to recognize several of the individuals who have in some way have helped and encouraged my
work with mosasaurs. I thank Gordon Bell, Pete Bussen, Kenneth Carpenter, Donald
Hattin, John Jagt, Greg Liggett, Johan Lindgren, Theagarten Lingham-Soliar, Earl Manning,
Larry Martin, James Martin, Judy Massare, Dan Merriam, Eric Mulder, Elizabeth Nicholls,
David Parris, Dale Russell, Bruce Schumacher, David Schwimmer, Kenshu Shimada, J. D.
Stewart, Glenn Storrs, Dan Varner and Richard Zakrzewski.
References (Not cited in the original article):
Bardet, N. and J. W. M. Jagt. 1996. Mosasaurus hoffmanni, le
"Grand Animal fossile des Carrieres de Maestricht": deux sicles d’histoire,
Bulletin de Museum d’Histoire naturelle, Paris, 4th Series, 18, Section C, No. 4, pp.
569-593. (in French, with a nice series of plates of the first discovered
mosasaur)
Bell, G. L. 1997. A phylogenetic revision of North American and Adriatic
Mosasauroidea. In Callaway J. M. and Nicholls, E. L., eds, Ancient Marine Reptiles,
Academic Press, pp. 293-332.
Bell, G. L. and J. P. VonLoh. 1998. New Records of Turonian Mosasauroids from
the Western United States. Fossil Vertebrates of the Niobrara Formation in South Dakota,
Dakoterra 5:15-28.
Cope, E. D. 1872. On the
geology and paleontology of the Cretaceous strata of Kansas. Preliminary Report of the
United States Geological Survey of Montana and Portions of the Adjacent Territories, Part
III - Paleontology, pp. 318-349.
Carpenter, K. 1990. Upward Continuity of the Niobrara Fauna with the Pierre
Shale Fauna. In S. C. Bennett, ed., Niobrara Chalk Excursion Guidebook, The University of
Kansas Museum of Natural History and the Kansas Geological Survey, pp. 73-81.
Everhart, M. J. 2001. Revisions to the
Biostratigraphy of the Mosasauridae (Squamata) in the Smoky Hill Chalk Member of the
Niobrara Chalk (Late Cretaceous) of Kansas. Kansas Acad. Sci. Trans 104(1-2):56-75.
Everhart, M. J. 2002. New data on cranial measurements and
body length of the mosasaur, Tylosaurus nepaeolicus (Squamata; Mosasauridae), from
the Niobrara Formation of western Kansas. Kansas Academy of Science, Transactions
105(1-2):33-43.
Everhart, M. J. 2002. New data on plesiosaur remains found as stomach contents
of a Tylosaurus proriger (Squamata; Mosasauridae) from the Niobrara Formation of
western Kansas. (Abstract) Kansas Academy of Science, Transactions
Everhart, M. J. 2002. Remains of immature mosasaurs (Squamata; Mosasauridae)
from the Niobrara Chalk (Late Cretaceous) argue against nearshore nurseries. (Abstract)
Journal of Vertebrate Paleontology 22(suppl. to 3):52A.
Everhart, M. J. 2004. Late Cretaceous
interaction between predators and prey. Evidence of feeding by two species of shark on a
mosasaur. PalArch, vertebrate palaeontology series 1(1):1-7.
Everhart, M. J. 2004. Plesiosaurs as the food of mosasaurs; new data on the stomach
contents of a Tylosaurus proriger (Squamata; Mosasauridae) from the Niobrara
Formation of western Kansas. The Mosasaur 7:41-46.
Everhart, M. J. 2005. Rapid evolution, diversification and distribution of
mosasaurs (Reptilia; Squamata) prior to the K-T Boundary. Tate 2005 11th Annual Symposium
in Paleontology and Geology, Casper, WY, p. 16-27 (not peer-reviewed).
Everhart, M. J. 2005. Oceans of Kansas - A Natural History of the Western Interior Sea.
Indiana University Press, 320 pp.
Everhart, M. J. 2005. Earliest record
of the genus Tylosaurus (Squamata; Mosasauridae) from the Fort Hays Limestone
(Lower Coniacian) of western Kansas. Kansas Academy of Science,
Transactions 108(3/4), p. 149-155.
Everhart, M. J. 2005. Tylosaurus kansasensis, a new species of
tylosaurine (Squamata: Mosasauridae) from the Niobrara Chalk of western Kansas, U.S.A.
Netherlands Journal of Geosciences / Geologie en Mijnbouw, 84(3), p. 231-240.
Gilmore, C. W. 1912. A new Mosasauroid Reptile from the Cretaceous of Alabama.
Proceedings U.S. National Museum, 40(1870):489-484.
Goldfuss, A. 1845. Der Schädelbau des Mosasaurus,
durch Beschreibung einer neuen Art dieser Gattung erläutert, Nova Acta Academa Ceasar
Leopoldino-Carolinae Germanicae Natura Curiosorum 21:1-28.
Hattin, D. E. 1982. Stratigraphy and depositional environment of Smoky Hill
Chalk Member, Niobrara Chalk (upper Cretaceous) of the type area, western Kansas. Kansas
Geol. Survey Bull. 225, 108 pp.
Jagt, J. W. M. 1995. Mosasauriers in het Natuurhistorisch Museum Maastricht
Opnieuw onder de loep Genomen Genomen (Mosasaurs in the Museum of Natural History
Maastricht - a review). Natuurhistorisch Maandblad, 84(2):46-59.
Liggett, G. A., S. C. Bennett, K. Shimada and J. Huenergarde. 1997. A Late
Cretaceous (Cenomanian) fauna in Russell county, KS. Kansas Acad. Sci. Trans. Abstracts.
16:26.
Lindgren, J. 1998. Early Campanian mosasaurs (Reptilia, Mosasauridae) from the
Kristianstad Basin, southern Sweden, Examensarbete I Geologi vid Lunds Universitet,
98:1-25.
Lindgren, J., Caldwell, M.W. and Jagt, J.W.M. 2008. New data on the postcranial anatomy
of the California mosasaur Plotosaurus bennisoni (Camp, 1942) (Upper Cretaceous:
Maastrichtian), and the taxonomic status of P. tuckeri (Camp, 1942). Journal of
Vertebrate Paleontology 28(4):1043-1054.
Lindgren, J. and M. J. Everhart. 2000. Remarks on two problematic mosasaur
specimens from the Smoky Hill Chalk (late Cretaceous) of Kansas. Kansas Acad. Sci. Trans.
19(abstracts):32.
Lingham-Soliar, T. 1995. Anatomy and functional morphology of the largest
marine reptile known, Mosasaurus hoffmanni (Mosasauridae, Reptilia) from the
Upper Cretaceous, Upper Maastrichtian of the Netherlands, Phil. Trans. R. Soc. Lond. B.
347:155-180.
Lingham-Soliar, T. 1998. A new mosasaur Pluridens walkeri from the Upper
Cretaceous Maastrichtian of the Lullemmeden Basin, southwest Niger. Journal of Vertebrate
Paleontology, 18(4):709-717.
Lingham-Soliar, T. 1999. What Happened 65 Million Years Ago: The study of giant
marine reptiles throws new light on the last major mass extinction. Science Spectra, No.
17:20-29.
Martin, James E. and P. R. Bjork. 1987. Gastric residues associated with a
mosasaur from the Late Cretaceous (Campanian) Pierre Shale in South Dakota. Dakoterra,
3:68-72.
Martin, J. E., B. A. Schumacher, D. C. Parris, and B. Smith-Grandstaff, 1998.
Fossil vertebrates of the Niobrara Formation in South Dakota. Dakoterra 5:39-54.
Martin, L. D. and J. D. Stewart. 1977. The oldest (Turonian) mosasaurs from
Kansas. Journal of Paleontology, 51(5):973-975.
Martin, L. D. and B. M. Rothschild. 1989. Paleopathology and diving mosasaurs. American
Scientist, 77:460-467.
Massare, J. A. 1994. Swimming capabilities of Mesozoic marine reptiles: a
review. pp. 133-149, In Maddock, L., Bone, Q., and Rayner, J. M. V. (eds.), Mechanics and
Physiology of Animal Swimming, Cambridge University Press.
Mulder, E. W. A. and H. Mai. 1999. The oldest tylosaurine mosasaur (Reptilia,
Lacertilia) from the Late Cretaceous of Belgium: Hermann von Meyer (1860) revisited,
Geologie en Mijnbouw, 78:207-213.
Nicholls, E. L. 1988. Marine vertebrates of the Pembina Member of the Pierre Shale
(Campanian, Upper Cretaceous) of Manitoba and their significance to the biogeography.
Ph.D. dissertation, The University of Calgary, 317 pp.
Nicholls, E. L. 1988. The first record of the mosasaur Hainosaurus
(Reptilia: Lacertilia) from North America. Canadian Journal of Earth Sciences 25,
1564-1570.
Nicholls, E. L. and A. P. Russell. 1990. Paleobiogeography of the Cretaceous Western
Interior Seaway of North America: the vertebrate evidence. Palaeogeography,
Palaeoclimatology, Palaeoecology, 79:149-169.
Parris, D. C., B. S. Grandstaff, R. K. Denton, and S. G. Lucas. 1997. Type locality of Liodon
dyspelor (Reptilia: Mosasauridae). Proc. Acad. Nat. Sci. Phila. 147:193-203.
Russell, D. A.. 1967. Systematics and morphology of American mosasaurs. Peabody
Museum of Natural History, Yale University, Bulletin 23.
Russell, D. A.. 1970. The vertebrate fauna of the Selma Formation of Alabama, Part VII,
The Mosasaurs. Fieldiana, Geology Memoirs, Vol. 3, No. 7, Field Museum of Natural History.
Russell, Dale A. 1975. A new species of Globidens from South Dakota. Fieldiana
Geology, Volume 33, No. 13, Field Museum of Natural History
Russell, D. A. 1993. Vertebrates in the Western Interior Sea. pp. 665-680, In
Caldwell, W. G. E. and E. G. Kaufmann (eds.), Evolution of the Western Interior Basin,
Geological Association of Canada, Special Paper 39.
Schumacher, B. A.. 1993. Biostratigraphy of Mosasauridae (Squamata, Varanoidea)
from the Smoky Hill Chalk Member, Niobrara Chalk (Upper Cretaceous) of Western Kansas.
Unpub. Masters Thesis, Fort Hays State University, 68 pp.
Schumacher, B. A. and J. E. Martin. 1993. First definitive record of the mosasaur Tylosaurus
proriger from the Niobrara Formation (upper Cretaceous), South Dakota. Proc. S.D.
Acad. Sci., 72:235-240.
Schumacher, B. A. and D. W. Varner. 1996. Mosasaur caudal
anatomy. Journal of Vertebrate Paleontology 16(Supplement to 3):63A.
Schwimmer, D. R., J. D. Stewart, and G. D. Williams. 1997. Scavenging by Sharks of
the Genus Squalicorax in the Late Cretaceous of North America. PALAIOS,
12:71-83.
Sheldon, M. A. 1996. Stratigraphic Distribution of Mosasaurs in the Niobrara
Formation of Kansas, Paludicola 1:21-31. (The Rochester Institute of Vertebrate
Paleontology)
Sheldon, M. A. 1997. Ecological implications of mosasaur bone microstructure. pp. 333-354
In Calloway, J. M. and Nicholls, E. L., eds, Ancient Marine Reptiles, Academic Press.
Shimada, K. 1997. Paleoecological relationships of the
Late Cretaceous lamniform shark, Cretoxyrhina mantelli (Agassiz), Journal of
Paleontology 71(5): 926-933
Stewart, J. D. 1988. The stratigraphic distribution of Late Cretaceous Protosphyraena
in Kansas and Alabama. Geology, Paleontology and Biostratigraphy of Western Kansas. in
Nelson, M. E. (ed.), Articles in Honor of Myrl V. Walker, Fort Hays Studies, Third Series,
No.10, Science, pp. 80-94.
Stewart, J. D. 1990. Niobrara Formation vertebrate stratigraphy. pp. 19-30, In
Bennett, S. C., ed., Niobrara Chalk Excursion Guidebook, The University of Kansas Museum
of Natural History and the Kansas Geological Survey.
Stewart, J. D. and Bell, G. L. Jr. 1994. North America’s oldest mosasaurs are
teleosts, Contributions to Science (Natural History Museum of Los Angeles County, No. 441,
pp. 1-9.
VonLoh, J. P. and Bell, G. L. Jr. 1998. Fossil reptiles from the Late
Cretaceous Greenhorn Formation (Late Cenomanian-Middle Turonian) of the Black Hills
Region, South Dakota. Dakoterra, 5:28-38.
Williston, S. W. 1898. Mosasaurs. The University Geological Survey of Kansas, Volume IV,
Paleontology, Part V, pp. 81-347.
Williston, S. W. 1904. The relationships and habits of the mosasaurs. Journal
of Geology, 12(4):43-51.
Williston, S. W. 1914. Water Reptiles of the Past and Present. Chicago
Univ. Press. 251 pp.

{kind=link}
{kind=link}
{kind=link}